The views expressed in this paper are those of the writer(s) and are not necessarily those of the ARJ Editor or Answers in Genesis.
Abstract
Aside from the brain, the liver is the most complex organ in the body. This irreducibly complex structure has over 500 critical functions in humans. A vertebrate cannot survive without a liver. Invertebrates do not have a liver or anything close, and an enormous unbridgeable gap exists between invertebrates and vertebrates in this area. As many of its functions are required for life, structures and cells exist in invertebrates that perform some of liver functions. Furthermore, the liver in all mammalian species is close to identical, and no evidence exists of any evolution of the liver. The progenitor to mammal liver was an organ close to identical to the liver in modern humans. Likewise, some mammals have a gallbladder connected associated with the liver, while others lack this support system, clearly showing that the first gallbladder was a fully functional organ which included all of its support systems.
Keywords: liver, gallbladder, evolution of vertebrate organs, evolution of the liver, irreducible complexity and intelligent design.
Introduction
The liver is an organ found only in vertebrates from fish to primates, and in no other animals. Since many of its functions are essential for life, invertebrates use other organs and cells to perform some of the liver functions. These smaller, less-complex appearing structures are very different from livers, but are likewise irreducibly complex systems. Examples are the cephalopod’s digestive gland has some of the liver functions. Earthworms have chloragogen cells that have some liver functions. Furthermore, vertebrate livers all have very similar designs and functions. Even the fish liver anatomy and function is basically similar to all other vertebrate livers (Bruslé and Gonzàlez i Anadon 1996). The main difference between the fish liver and the mammalian liver is its morphological shape. The mammalian, and all other vertebrate livers conform to the shapes and arrangement of contiguous viscera and peritoneal bounds. The liver is typically the largest visceral organ in the body. Furthermore
The microarchitecture of the liver is very similar among mammalian species and best represented by the lobular concept, with the biggest difference present in the degree of connective tissue development in the portal tracts. (Kruepunga et al. 2019)
The substances it can detoxify also vary, depending on the animal’s diet. A common example is that humans are able to eat chocolate without problems, but dogs cannot; consequently, chocolate can kill a dog. For this reason, even normally harmless foods, along with alcohol and drugs, are not equally tolerated across all species.
Two basic types of livers exist, the tunneled parenchyma found only in mammals, and the net-like tubular glands in non-mammal vertebrates. Furthermore, no intermediate forms are found that could conceivably link these two. This raises the question: “How did this type of liver, [the mammalian], evolve from the liver of the lower vertebrates?” (Elias and Bengelsdorf 1952, 297). Parenchyma refers to the functional tissue as opposed to the stroma (structural, support, and vascular tissue). As Reynolds describes the difference, the mammalian liver is a “continuous mass of parenchymal cells tunneled by vessels through which venous blood flows on its way from the gut to the heart” (Klassen 1975; Reynolds 1977, 1). This unique design, called the hepatic portal system where blood leaving one capillary bed (in the gut) goes to another capillary bed (in the liver). Thus two capillary systems are joined by veins.
The Gallbladder
Another difference between livers is the gallbladder, which is located below the liver and used to store bile produced by the liver and released during meals to help digest fat. Another function of the gallbladder in humans is to reabsorb water, concentrating bile solution up to ten times. The gallbladder exists in humans and in most other mammals, but is lacking in a few mammal types, such as rats (Gorham and Ivy 1938). The dilemma is that the mouse, for example, has a gallbladder while its supposed evolutionary relative, the rat, is lacking this organ. One study sought to determine whether if in species lacking a gallbladder, the bile concentrating function was in the ducts (McMaster 1922, 127). The study found that not only is the gallbladder absent in rats, but also one of its important functions is absent, namely the concentration of bile. The research concluded that
the mouse secretes somewhat more bile per 100 gm. of body weight. This bile as it comes from the liver is but one-eighth as strong at least in pigment as rat bile, but the concentrating activity of the gallbladder is so great that the products yielded to the intestine may become not dissimilar. (McMaster 1922, 140).
In short, lack of a gall bladder is compensated for by production of more concentrated bile (Friman et al. 1990). Other examples of this dilemma are the gallbladder’s presence in cows, sheep, and goats but its absence in horses and deer. Although found in the hog and wild boar, it is absent in the closely related peccary pig (McMaster 1922, 127).
The problem is that “evolutionary biology claims that life came about by chance and laws of nature alone, so it must explain how transitional organisms survived as the liver gradually acquired these vital functions while macroevolution was taking place” (Glicksman 2016). Specifically, the problem for evolutionists is how could the systems used in invertebrates become incorporated into a new, very different organ that had functions no inveterate had? No one has proposed a viable theory and the problem is ignored.
Functions of the Liver
The liver has over 500 functions and each one is important for survival of vertebrates. Its work is done in functional units called lobules that are composed of hepatocytes arranged in a pattern to deliver the cell’s products into the blood stream (see fig. 1.). The extent of the liver factory is illustrated by the fact that close to one-million lobules exist, each one-millimeter in diameter (Olsen 2009, 101). These lobules function to detoxify various metabolites, synthesize proteins, regulate glycogen storage, and produce hormones and biochemicals required for digestion and growth. It also is involved in cycles (such as the Cori cycle), that impact the metabolism of carbohydrates, proteins, and fats for body energy and nutrient needs. Another function of the liver is to produce numerous enzymes that chemically break down drugs, hormones, and other substances to regulate the body’s chemical environment. Lastly, it produces several proteins including fibrinogen and prothrombin, both required for blood clotting. Without these two factors bleeding to death would be common. Examples that affect fibrinogen and prothrombin include the rare coagulation disorders (RCDs). On example is Factor X Deficiency. Factor X is produced in the liver (Greipp , Kyle, and Bowie 1981, 443; Spiliopoulos et al. 2019, 195). Another example is vitamin K deficiency, especially in infants, which causes a prothrombin deficiency (Shearer 2009, 49).

Fig. 1. The anatomy of the liver showing how the products of the hepatocytes enter the blood stream and the blood is processed by the hepatocytes. https://commons.wikimedia.org/wiki/File:2423_Microscopic_Anatomy_of_Liver.jpg.
Conversion of Ammonia into Urea
The liver converts the highly toxic by-product of protein metabolism, ammonia, into urea which is then expelled from the body through the kidneys. This conversion requires five specific enzymes in the urea cycle. If any one of these five enzymes is absent, life would be impossible because elevated ammonia blood levels are very toxic to the nervous system. One example is urea cycle disorder which produces profound results from toxic levels of ammonia including irreversible toxicity in newborns (Matsumoto 2019, 833). The problem is the urea cycle is missing a required enzyme that converts nitrogen waste into urea. Untreated, the high amounts of ammonia in the blood stream causes brain damage, coma and, eventually, death. Thus without the liver, ammonia and other toxic substances would end up in the blood, causing problems including hepatic encephalopathy. Even mild levels cause irritability, short attention span, anxiety, and insomnia. Higher levels cause lethargy, confusion, slurred speech, and are eventually fatal. Invertebrates must utilize a very different design to deal with any toxic byproduct of protein metabolism.
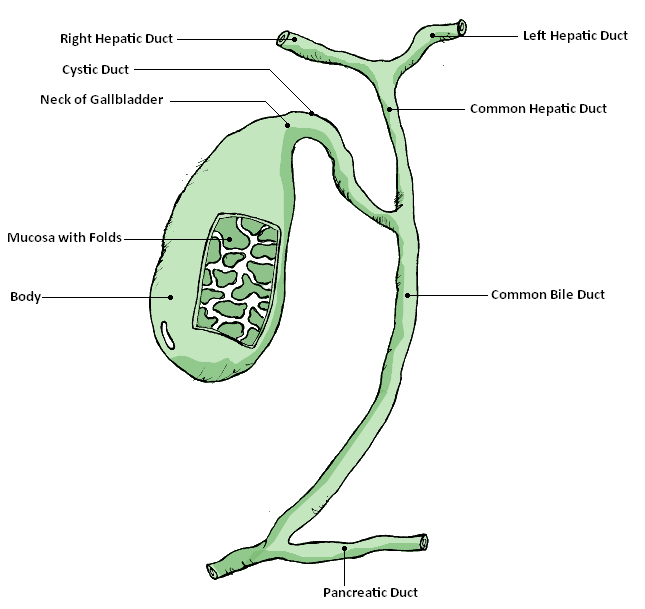
The liver also helps the body break down red blood cells and, using enzymes, chemically converts hemoglobin into bilirubin. During the transition from fetal to adult hemoglobin in newborns, the liver can be “overwhelmed” and unable keep up with the bilirubin production, resulting in neonatal jaundice. The reason why is a newborn’s liver is not fully developed. Consequently, it is less effective at processing bilirubin and removing it from the blood. At around two weeks, less bilirubin is produced and the liver is more effective at removing it from the body. For these reasons jaundice normally corrects itself without causing harm. The bilirubin is then combined with cholesterol and other chemicals to form bile. The bile moves through the common hepatic duct where it coalesces with the cystic duct from the gallbladder forming the common bile duct which then empties into the first part of the intestines called the duodenum (see fig. 2). In the intestine bile functions as an emulsifier to help the intestines absorb fats and, with it, fat-soluble vitamins including vitamin A for vision, vitamin E, an anti-oxidant, vitamin K for blood clotting, and vitamin D for bone strength and calcium metabolism (Housset et al., 2016).
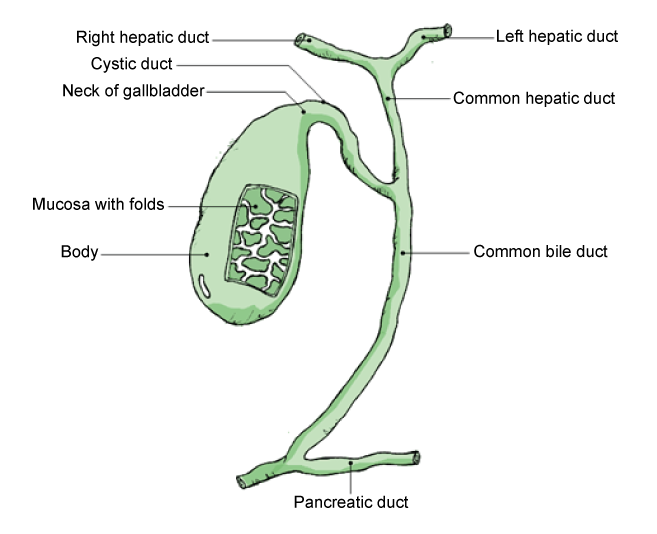
Fig. 2. The gallbladder and its duct system. From Wikimedia-Commons. Source: GallbladderAnatomy.png. Author: GallbladderAnatomy.png by LukesAnatomy (talk); conversion to SVG by Angelito7 (talk). https://upload.wikimedia.org/wikipedia/commons/6/61/GallbladderAnatomy.png.
Albumin
Over 60% of the liver’s entire protein output is the protein albumin. Albumin aids in transporting materials in the blood to where they are needed in the body (Rosenoer, Oratz, and Rothschild 2014). This includes the minerals calcium and magnesium, plus certain lipids, and steroid hormones including estrogen, progesterone, and testosterone.
It also maintains the blood volume by keeping enough water in circulation by preventing fluids from leaking into other tissues. As blood enters the arterial side of the capillary under pressure, it moves to the venous side, resulting in water loss. The water is pushed out of the circulation through the pores within the capillary walls and into the interstitial fluid. Albumin is a large protein that cannot pass through the capillary walls.
As a large very polar molecule it requires a huge number of surrounding water molecules to keep it in solution, thus exerting a reverse osmotic pressure called oncotic pressure. Albumin (and associated sodium) increases the osmotic pressure in the plasma, which draws water back from the interstitial fluid into the capillaries. Water loss is therefore directly related to the albumin level in the blood. If the liver doesn’t produce enough albumin, more water tends to move out of circulation and into the interstitial fluid, reducing blood volume and causing blood pressure to drop. Low albumin levels increase abdominal fluid buildup and leg edema. The liver has an intricate system to measure the osmotic pressure of the blood, producing albumin when the osmotic pressure drops to increase plasma fluid retention. This system is only necessary in animals with closed circulatory systems.
The Complement Immune System
The liver produces a set of 30 different proteins that help the immune cells fight infection. Complement proteins are released by the liver into the blood, but are inactive until they encounter microbial infections (Ross 2014). This encounter triggers them to increase inflammation in the infection area that kills the microbes, and also to attach to microbes, attracting immune cells that ingest the microbes in a process called opsonization. If liver problems produce less complement proteins the body is more susceptible to a wide variety of infections, both viral and bacterial, including E. coli 0157, Salmonella, and Staphylococcus aureus. Another critical function of the liver is detoxification achieved by chemically converting toxins into waste products. An example is alcohol detoxified in the liver by liver peroxisomes in liver cells. Detoxifying alcohol and certain other harmful compounds is achieved by transferring hydrogen from the toxin to oxygen, a process termed oxidation. Acetaminophen in Tylenol detoxification is also an important task of the liver, and if the build-up is greater than the liver can handle, the acetaminophen can damage the liver.
Liver Evolution
As briefly described above, evolutionary biologists are unable to even imagine how an ultra-complex organ like the liver could have ever come into being; although how it actually functions to keep the body alive is fairly well understood. As noted above, our earliest ancestors could not survive without a means to carry out all of the functions of the liver which in invertebrates are done by specific cell groups not part of a liver.
The most common reason evolutionists provide for the lack of evidence bridging the chasm between the non-liver and liver is the fact that soft tissue is normally not preserved in the fossil record. However, this claim ignores the fact that thousands of so-called “living fossils” exist which are believed to be anatomically close to their claimed multiple millions-of-years-old designs. These “living fossils” should display evidence of less-evolved livers, but do not. In other words, if the evolution of the liver occurred, evidence of it should exist in “living fossil” animals. Lower organisms don’t require certain liver functions like blood clotting or osmoregulation via albumin, but do require some functions the vertebrate liver performs, such as detoxification. One system is Metallothioneins used in aquatic invertebrates to facilitate metal detoxification of mercury, silver, cadmium and other metal toxins (Amiard et al. 2006, 160). Metallothioneins also function to help protect the body from free radical damage. Metallothioneins are non-enzymatic proteins that function by their cysteine thiol groups binding to certain heavy metals, detoxifying and excreting them (Vogt and Quinitio 1994).
Most references on organ evolution completely ignore the difficulty of the evolution of the liver. This includes the standard text in this area by Schmidt- Rhaesa (2007; Torday, Blackstone, and Rehan 2019). Downard and Wheat’s (2020) mammoth 828-page answer to Intelligent Design likewise totally ignores the topic. Olson (2009, 100–102) mentions the liver four times and has an entire section on it in his book, yet never mentions its evolution. He defines evolution as “a comprehensive and well-established scientific explanation, based on natural processes, of the fact of biological evolution.” Evolution is so fully proved that, he adds, “there is no longer a compelling reason to keep testing or looking for examples of evolution.”
Yet in his 211-page over-sized book, excepting the brain and birth canal, no hint of the evolution of any of the human body organs and structures was presented. This is true even in the earliest books on organ evolution such as Wilder (1909). One study concluded that the Neanderthal rib-cage evolved to accommodate a larger liver, which the authors speculate had evolved in Larmarkian fashion due to changes in the Neanderthal diet toward more meat protein (Ben-Dor, Gopher, and Barkai 2016). No viable origin of the liver is addressed in this work on Neanderthals.
Since fish, as vertebrates, also have a liver, the assumption is made that all mammals have a liver because they evolved from fish. Therefore, mammal livers should all be genetically very similar. But this is not what has been found. The few articles that cover the liver evolution topic state only that mammals all evolved from some hypothetical first mammal; thus all mammals have similar liver systems. Going back farther, they imply that the early hypothetical vertebrate precursor had a liver, an assumption that explains the liver’s existence in all vertebrates. Other than this assumption they do not address how the first liver evolved.
One study compared 4,000 human genes with nearly identical counterparts, known as homologous genes, in mouse liver cells (Odom et al. 2007). The genetic research on gene regulation of mice and human liver cells found that “the number of genes with identical regulation in both species was very, very small.” This is in stark contrast to the expectation of evolutionists that there would be very little difference based on similar evolutionary origins and close to identical functions. Specifically, they found that
Despite the conserved function of these factors, from 41% to 89% of their binding events seem to be species specific. When the same protein binds the promoters of orthologous genes, approximately two-thirds of the binding sites do not align. (Odom et al. 2007)
The researchers concluded, personifying evolution, that “evolution has discovered several different ways to make [a] liver from the same building blocks” (Trafton 2007; emphasis added). They used mice as a human biology model because of the homologous function of liver and basic biological processes of mice are assumed to be essentially the same as humans.
Summary
No evidence of the evolution of the liver was found to bridge the enormous gap between the liver and comparable systems in invertebrates such as the Metallothioneins used in aquatic invertebrates. Although some liver functions exist in invertebrates, their design is very different than vertebrates. These proteins function for the life forms needs, but would not meet the requirement of vertebrates. Consequently, a very different complex system is employed in vertebrates called a liver. An enormous problem is explaining how this irreducibly complex system evolved from the very different systems used in invertebrates, given the fact that most of its functions are required for vertebrate life (DeBenedictis, 2014, chapter 14).
Acknowledgements
Acknowledgements include Kirk Ktoth, David V. Bassett, and especially Howard Glicksman MD for their help and feedback.
References
Amiard, J.-C., C. Amiard-Triquet, S. Barka, J. Pellerin, and P. S. Rainbow. 2006. “Metallothioneins in Aquatic Invertebrates: Their Role in Metal Detoxification and Their Use as Biomarkers.” Aquatic Toxicology 76, no. 2 (10 February): 160–202.
Ben-Dor, Miki, Avi Gopher, and Ran Barkai. 2016. “Neandertals’ Large Lower Thorax May Represent Adaptation to High Protein Diet.” American Journal of Physical Anthropology 160, no. 3 (June): 367–378.
Bruslé, Jacques, and Gemma Gonzàlez i Anadon. 1996. “The Structure and Function of Fish Liver.” In Fish Morphology, edited by J. S. Datta Munshi and Hiran M. Dutta. CRC Press, Boca Raton, Florida.
DeBenedictis, Albert. 2014. Evolution or Creation? A Comparison of the Arguments. 3rd ed. Xlibris: Bloomington, Indiana.
Downard, James, and Jackson Wheat. 2020. The Rocks Were There: Straight Science Answers to Some Bent Creationist Questions. Vol. 1. The Troubles in Paradise Project. www.tortucan.wordpress.com.
Elias, H., and H. Bengelsdorf. 1952. “The Structure of the Liver of Vertebrates.” Acta Anatomica 14, no. 4 (July 1): 297–337.
Friman, S., G. Ràdberg, I. Bosaeus, and J. Svanik. 1990. “Hepatobiliary Compensation for the Loss of Gallbladder Function After Cholecystectomy. An Experimental Study in the Cat.” Scandinavian Journal of Gastroenterology 25, no. 3 (March): 307–314.
Glicksman, Howard. 2016. “Liver Function and Its Effects.” Evolution News, June 11. https://evolutionnews.org/2016/06/liver_function/.
Gorham, Frank W. and Andrew Conway Ivy. 1938. “General Function of the Gallbladder from the Evolutionary Standpoint.” Field Museum of Natural History, Zoology Series 22, no. 3 (June 21): 159–213.
Greipp, P. R., R. A. Kyle, and E. J. Bowie. 1981. “Factor-X Deficiency in Amyloidosis: A Critical Review.” American Journal of Hematology 11, no. 4 (December): 443–450.
Housset, Chantal, Yues Chrétien, Dominique Debray, and Nicolas Chignard. 2016. “Functions of the Gallbladder.” Comprehensive Physiology 6, no. 3 (June 13): 1549–1578.
Klassen, C. D. 1975. “Biliary Excretion of Xenobiotics.” CRC Critical Reviews of Toxicology 4, no. 1 (October): 1–29.
Kruepunga, Nutmethee, Theodorus B. M. Hakvoort, Jill P. J. M. Hikspoors, S. Eleonore Köhler, and Wouter H. Lamers. 2019. “Anatomy of Rodent and Human Livers: What are the Differences?” Biochimica et Biophysica Acta Molecular Basis of Disease 1865, no. 5 (May 1): 869–878.
McMaster, P. D. 1922. “Do Species Lacking a Gallbladder Possess its Functional Equivalent?” Journal of Experimental Medicine 35, no. 2 (January): 127–140.
Matsumoto, Shirou, Johannes Haberle, Jun Kido, Hiroshi Mitsubuchi, Fumio Endo, and Kimitoshi Nakamura. 2019. “Urea Cycle Disorders—Update.” Journal of Human Genetics 64, no. 9 (September): 833–847.
Odom, Duncan T., Robin D. Dowell, Elizabeth S. Jacobsen, William Gordon, Timothy W. Danford, Kenzie D. MacIsaac, P. Alexander Rolfe, et al. 2007. “Tissue-Specific Transcriptional Regulation has Diverged Significantly Between Human and Mouse.” Nature Genetics 39, no. 6 (21 May): 730–732.
Olsen, Bruce D. 2009. Understanding Human Anatomy Through Evolution. 2nd ed. Morrisville, North Carolina: Lulu Press.
Reynolds, Edward S. 1977. “Environmental Aspects of Injury and Disease: Liver and Bile Ducts.” Environmental Health Perspectives 20 (October): 1–13.
Rosenoer, Victor M., Murray Oratz, and Marcus A. Rothschild. 2014. Albumin: Structure, Function and Uses. New York, New York: Pergamon Press.
Ross, Gordon D. 2014. Immunobiology of the Complement System. New York, New York: Academic Press.
Schmidt-Rhaesa, Andreas. 2007. The Evolution of Organ Systems. New York, New York: Oxford University Press.
Shearer, Martin J. 2009. “Vitamin K Deficiency Bleeding (VKDB) in Early Infancy.” Blood Reviews 23, no. 2 (March): 49–59.
Spiliopoulos, Dimitrios and Rexan A. Kadir. 2019. “Congenital Factor X Deficiency in Women: A Systematic Review of the Literature.” Haemophilia 25. no. 2 (March): 195–294.
Torday, John S., Neil W. Blackstone, and Virender K. Rehan. 2019. Evidence-Based Evolutionary Medicine. Hoboken, New Jersey: Wiley-Blackwell.
Trafton, Anne. 2007. “Mice and Men Make Livers Differently.” MIT News, May 21. https://news.mit.edu/2007/evolution-0521.
Vogt, Günter, and Emilia T. Quinitio, 1994. “Accumulation and Excretion of Metal Granules in the Prawn, Penaeus monodon, Exposed to Water-Borne Copper, Lead, Iron and Calcium.” Aquatic Toxicology 28, nos. 3–4 (April): 223–241.
Wilder, Harris Hawthorne. 1909. History of the Human Body. New York, New York: Henry Holt.










{kind=link}
{kind=link}