
The views expressed in this paper are those of the writer(s) and are not necessarily those of the ARJ Editor or Answers in Genesis.
Abstract
Thick sequences of upper Cambrian (Furongian) microbialites have now been documented from North America, China, Korea, Russia, Australia, Iran, Argentina, Kyrgyzstan, and Antarctica. Since many creationists believe that Cambrian rocks were deposited in the Noahic Flood, then this would naturally imply that these thick sequences grew in less than one year. In this paper, the upper Cambrian microbialites from North America, China, and Korea are described in detail, and subsequently interpreted as in situ environments representative of time frames greater than that of the year-long Flood. This paper suggests that Cambrian microbialite assemblages grew in place during antediluvian time, and that the Great Unconformity is but one of many formed during Creation Week.
Keywords: Upper Cambrian; Stromatolites; pre-Flood; Flood, post-Flood; Precambrian; Great Unconformity; Creationism; Microbialites
Introduction
Most creationists believe that the Flood of Noah was global in scope, and that consequently, plenty of geological evidence should support this belief including the approximate location of the pre-Flood/Flood and Flood/post-Flood boundaries. Moreover, since most creationists interpret the geological events associated with the Flood in terms of “normal, natural processes,” then this geological evidence should be subject to scientific enquiry. Since the normal rules of scientific investigation apply, most creationists believe that the evidence for these boundaries should, therefore, be detectable and measurable.
Many creationists are actively engaged in finding the Flood/post-Flood boundary, but little work has been done on the pre-Flood/Flood boundary, primarily because most creationists consider the Precambrian-Cambrian contact geologically attractive (Austin and Wise 1994; Dickens 2017; Dickens and Snelling 2008; Snelling 2009). Arguments supporting a pre-Flood/Flood boundary at the Precambrian-Cambrian boundary are quite persuasive and make sense given the kinds of processes one might assume were at work during this period. Perhaps the most persuasive argument is the widespread existence of a surface of erosion called “the Great Unconformity.” This surface of erosion occurs at many localities around the world, and its existence is supported by both creationists and secularists alike (Austin and Wise 1994; Dickens 2017; Dickens and Snelling 2008; Peters and Gaines 2012). Another persuasive argument points to the great disparity that exists between the fossil record as it appears both below and above this boundary. Fossils, especially metazoans, are virtually absent in Precambrian rocks, while abundant in those rocks that belong to the Cambrian. Catastrophic burial during the Flood seems a robust explanation for the existence of exceptionally fossilized fauna all over the world. For most creationists then, the Precambrian-Cambrian Flood boundary is an obvious choice that needs no further consideration. This paper identifies some very real challenges to this assumption.
Perhaps the greatest challenge to the Precambrian-Cambrian pre-Flood/Flood boundary is the existence of in situ upper Cambrian (Furongian)1 “reefs” that were constructed by microbialite/sponge communities. These “reefs” are found in many places in North America, China, Korea, Russia, Australia, Iran, Argentina, Kyrgyzstan, and Antarctica (Hong et al. 2012; Lee, Sung, and Chough 2015). In this paper, I will provide a detailed description of such “reefs” in Utah and add to that a less detailed examination of other “reefal” systems throughout North America. I will then provide a synopsis of most of the upper Cambrian “reefal” systems that exist in Korea and China. In follow-up work, I hope to similarly document other Cambrian “reefs” from other continents.
Microbialite and “Reef” Definitions
For the purpose of this paper, the microbialite definition of Burne and Moore (1987, 241–242) will be used: “Microbialites are organosedimentary deposits that have accreted as a result of a benthic microbial community trapping and binding detrital sediment and/or forming the locus of mineral precipitation.” A microbialite is technically the umbrella term used for three basic fabrics that can be found within the overall macro-structure (the general external shape). These fabrics are given names: stromatolites, thrombolites, and dendrolites. Stromatolites have a concave down laminar fabric that is very distinctive (fig. 1A). Thrombolites have a clotted fabric without any laminations (fig. 1B). Dendrolites have a branching, shrub-like fabric (fig. 1C). The macro-structure, although varying from domed to column-like, can incorporate any of these three fabrics. You really must “crack them open” to find out what’s inside!
Microbialites accrete at a laminar to sub-laminar level using different processes. The simplest is the purely mechanical interaction whereby grains of sediment get stuck in the sticky, extracellular polysaccharide sheaths (EPS) of cyanobacterial communities. These kinds of microbialites often take on a stromatolitic texture. Sometimes, the cyanobacteria can cause the chemistry in the water around them to change. When this happens, micrite precipitates directly out of the water column and is deposited around the microbial community as carbonate mud. This kind of mechanism can produce either stromatolites or thrombolites. Calcimicrobes are cyanobacteria that actually become calcified. In other words, the organism or community of organisms hardens into calcite. A community of calcified filamentous Epiphyton, for example, can look like a small calcified “shrub” (fig. 1C and D). These forms almost never produce stromatolites, but often produce thrombolites and/or dendrolites (Kennard and James 1986) (fig. 1C).
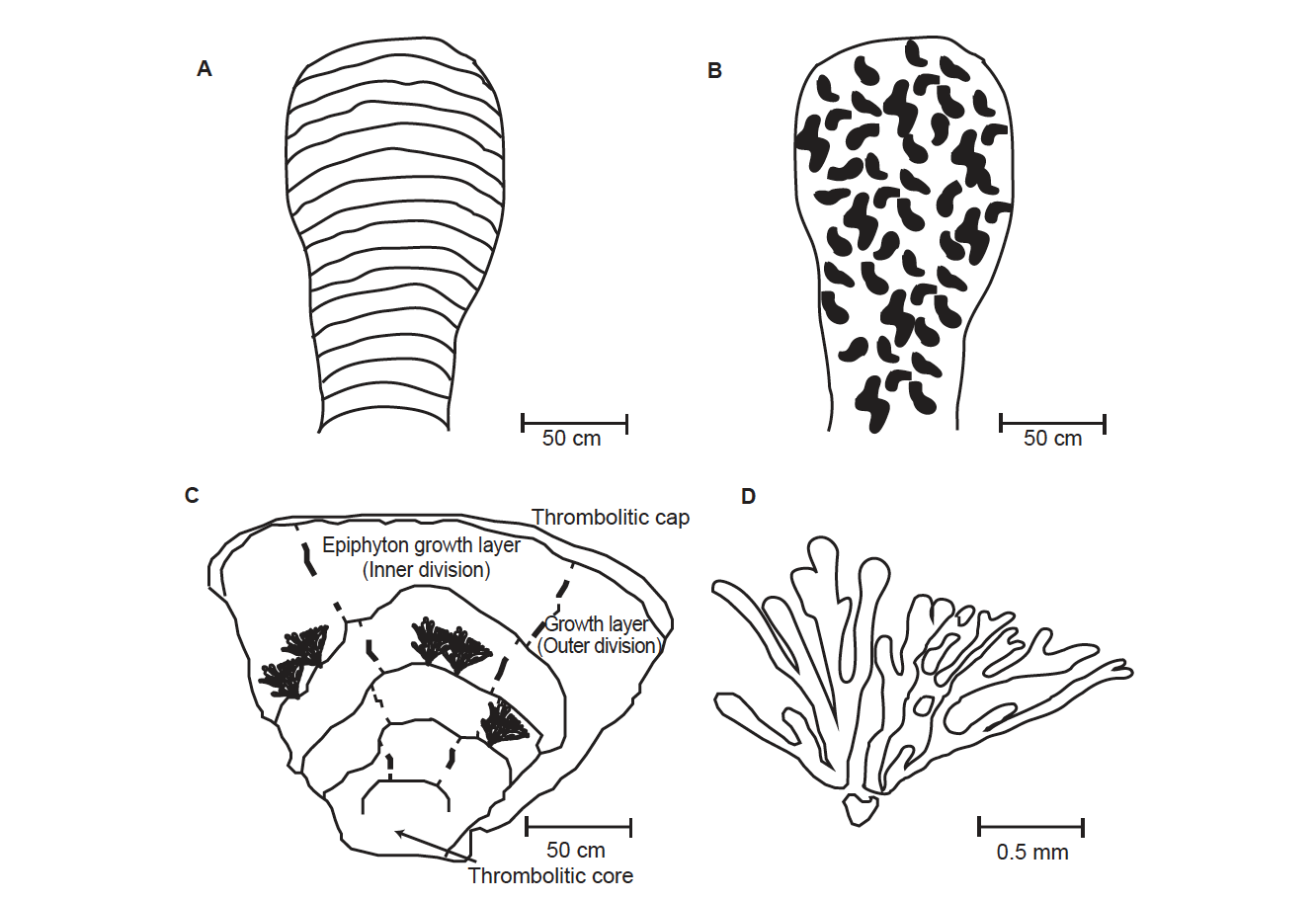
Fig. 1. A. Stromatolite. B. Thrombolite. C. Dendrolite. This dendrolite is actually composed of Epiphyton “shrubs.” These “shrubs” have been enlarged for clarity, but the actual size of a single “shrub” is about 1–2 mm across. Only a few “shrubs” are shown. D. A single Epiphyton “shrub.” Adapted from Woo and Chough (2010).
Cambrian “reefs” were chiefly constructed by non-calcifying, cyanobacteria, and/or calcifying filamentous cyanobacteria such as Epiphyton and Renalcis, along with various kinds of siliceous sponges, or sponge-like organisms. Nevertheless, workers consistently apply the term “reef,” and rightly I might add, even to the most basic of these structures— the humble microbialite. Yes, these were simple structures, but unlike later “mud-mounds,” these early microbial “reefs” were organically cemented together during construction, were durable, and were flanked by unconsolidated carbonate sediments. Given these characteristics, and given the substantial topographic relief that some of these “reefs” exhibited, I only think it appropriate to adopt the definition of “reef” put forward by Wood (1999): “A reef is a discrete carbonate structure formed by in situ or bound organic components that develops topographic relief upon the sea floor.” Given this definition, the term “reef” will, from this point forward, always appear without quotation marks. Of course, these early Cambrian reefs must be differentiated from the massive structures that most of us recognize as reefs today. The two are clearly different.
Investigations of Cambrian Microbialites
Cambrian Microbialites from Utah
The 550 m thick Notch Peak Formation of western Utah sits stratigraphically towards the top of an approximately 2600 m thick sequence of carbonates, and his been divided into three mappable members: the Hellnmaria, Red Tops, and Lava Dam (fig. 2). All three members are traceable from within the House and Confusion Ranges in western-central Utah to the Wah Wah Mountains in the south (Hintze, Taylor, and Miller 1988).
The area in and around western Utah has been extensively mapped by dozens of geologists over the past 120 years (Hintze and Davis 2003). Dozens of stratigraphic maps and geologic correlations have been painstakingly wrought from the exceptionally well-exposed rocks of western Utah. These maps and correlations have been further correlated by other workers to other formations existing throughout western and central North America, providing a very high-resolution stratigraphic picture that is quite comprehensive (Miller, Evans, and Dattilo 2012a; Miller, Loch, and Taylor 2012b).
The sequence specific to this paper has also received excellent stratigraphic coverage, with many comprehensive stratigraphic maps existing in the literature. Hintze and Davis (2003) constructed some of these maps, with part of their comprehensive, low resolution stratigraphic column from page 33 being partially reproduced here in fig. 3. In the original paper, this figure shows a stacked series of continuous strata starting with crystalline basement rock and extending all the way through to the Permian. Mesozoic and Cenozoic deposits also exist atop the Paleozoic sequence but are broken by large gaps of missing sediment. Although the gneiss bedrock is not exposed in the House Range area, it is exposed in the Mineral Mountains area about 50 mi to the southeast. Hintze and Davis (2003) also use borehole cores, aeromagnetic data, gravity data, earthquake seismicity data, and data from the Deep Reflection Seismic Survey, to construct a number of sub-surface maps that conclusively show that western Utah is underlain with crystalline basement (see their map on page 214 as an example). The Cambrian Prospect Mountain Quartzite is exposed at a number of sites in and around the House Range, and correlates to the Tintic Quartzite in central Utah, and the Geertsen Canyon Quartzite in northern Utah, all of which correlate to the Tapeats Sandstone in Grand Canyon area (Yonkee et al. 2014) which overlies the Great Unconformity.
The upper Cambrian (Furongian) Hellnmaria microbialites occur in eleven distinct microbialbearing beds that span the upper 154 m of the Hellnmaria Member within the Notch Peak Formation (Coulson and Brand 2016; Coulson, Brand, and Chadwick 2016; Hintze, Taylor, and Miller 1988) (figs. 2 and 3). Brand, Phillips, and Chadwick (2012) were able to trace some of these upper Hellnmaria microbialites to the Drum Mountains in the north and the Wah Wah mountains in the south, providing a total areal distribution of over 2,600 km2. Based on the work of others, it is likely that the total areal distribution for associated upper Cambrian microbialites in this area reaches to several tens of thousands of square kilometers (Hintze, Taylor, and Miller 1988; Miller et al. 2003; Miller, Evans, and Dattilo 2012a; Miller, Loch, and Taylor 2012b; Shapiro and Awramik 2000).
Although 11 beds occur within the Hellnmaria Member, my research discusses only two of them in detail. Bed 9 (fig. 2) is a 5–14 m thick stromatolitic unit that exhibits a change in morphology as seen in vertical cross-section. Forms change from round at the bottom of the bed to elongate in the middle of the bed and then back to round again at the top (fig. 2). Remarkably, each of these changing morphologies can be distinguished at seven outcrop locations, with the stromatolites in the elongate layer exhibiting a consistent 140°/320° bearing. At most of the outcrop locations, coalescing round forms are found both beneath and above the strongly elongate layer. These observations led me to hypothesize the existence of a bidirectional hydrodynamic system that was chiefly at work during the deposition of the elongate layer (Coulson, Brand, and Chadwick 2016). As a result, coalescence occurred parallel to flow constructing linear groups of laterally-linked forms. Strongly elongate structures naturally followed. The round forms at the top of the bed resulted from the removal of this bi-directional hydrodynamic system.
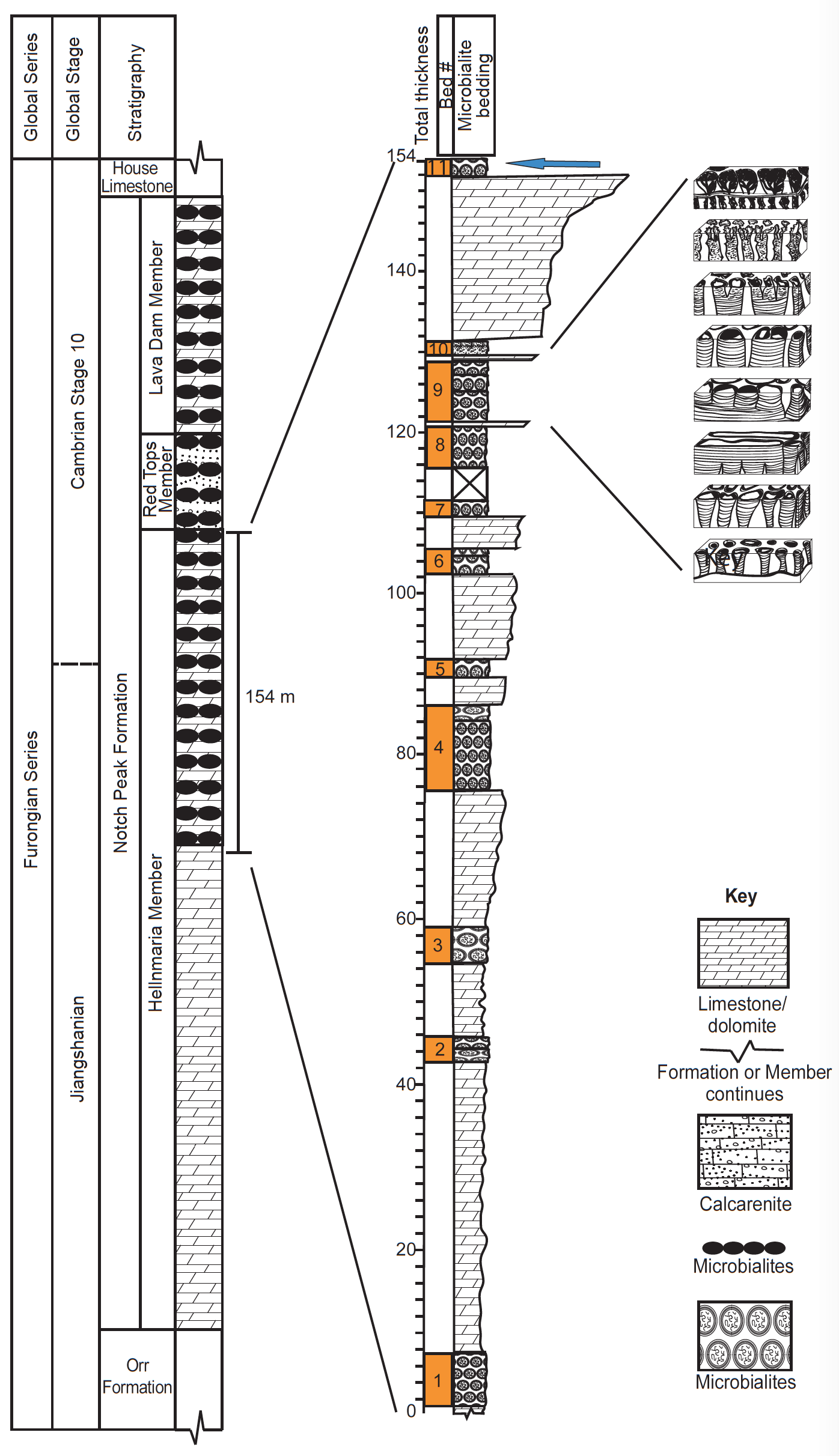
Fig. 2. Stratigraphic column of the Notch Peak Formation, adapted and simplified from Hintze, Taylor, and Miller (1988). Tapered Limestone/dolostone sections communicate a general coarsening upward trend. Arrow indicates bed 11. Bed 9 is expanded to the right. Notice individual round forms changing up-section into coalesced elongate forms, then changing back to “raggedy” round forms at the top. Note multiple layers of microbialites in both the Red Tops Member and the Lava Dam Member (not discussed in this paper). Figure modified and used with permission from SEPM, Coulson and Brand (2016).
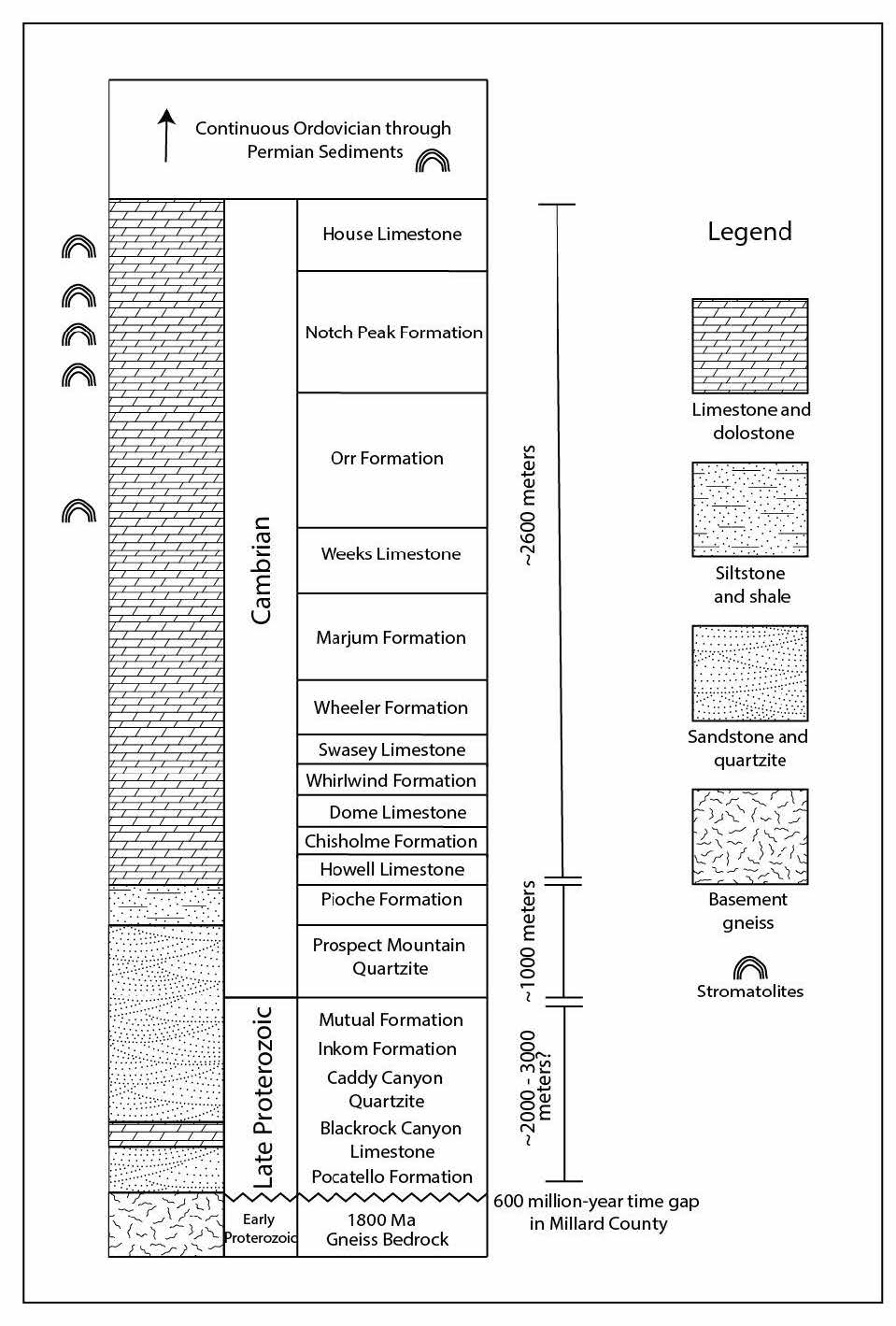
Fig. 3. Low resolution stratigraphic column adapted from Hintze and Davis (2003). Note that the Cambrian strata, including the Notch Peak Formation (this paper), are sitting on crystalline basement rocks. The middle-upper Cambrian has multiple microbialite assemblages. This paper only discusses the 11 microbialite horizons in the upper part of the Notch Peak Formation (Hellnmaria Member). These microbialite assemblages continue through into Ordovician strata as well.

Fig. 4. Macro and meso-scale features of Bed 11. Abbreviations: S = stromatolite; IS = interspace; TC = Thrombolitic Core; MS = mini-stromatolites; MSC = mini-stromatolite core. A. Large, round microbialites are tightly packed with respect to each other, a feature that is common throughout the entire bed. B. An in situ microbialite that was pulled directly from its growth position in bed 11 (see fig. 9D). Notice the concave-down laminations middle to bottom, as well as thrombolitic core, middle to top. These thrombolitic cores contain mini-stromatolites (the black square in B is expanded in C). D. An in situ microbialite that was also pulled directly from its growth position in bed 11 (see fig. 9D). The microbialite was taken back to the lab and cut longitudinally through the medial plane. Again, notice concave-down laminations. Centimeter scale at bottom of image. E. A similar microbialite as in D, but this time cut transversely, revealing the maze-like structure of the mini-stromatolites. F. The interspace between these microbialites is full of trilobite hash, mollusks, and limestone intraclasts. Figure modified and used with permission from SEPM, Figure modified and used with permission from SEPM, Coulson and Brand (2016).
Bed 11 (fig. 2) is a 1–3 m thick stromatolitic unit that contains a tightly packed field of round to subround forms (fig. 4). Mesoscale fabric is best described as stromatolitic, but many forms have a large central thrombolitic core composed of mini-stromatolites (figs. 4B, C, and E). Diameters vary from about 40–70 cm, and due to the fissile nature of the overlying strata, are very well exposed in plan-view at multiple locations over an area of about 20 km2. Heights vary from 20 cm to about 70 cm and in cross-section widen slightly towards the top (figs. 4B and D). During my time at the Utah research site, I saw literally hundreds of stromatolites, all of which were upright and in apparent growth position.
Within many of the Bed 11 stromatolites I found sponge-spicule networks (fig. 5). These networks represent the remains of siliceous sponges that calcified before postmortem decay, leaving the isolated spicules “floating” in what may have originally been a fleshy matrix (Coulson and Brand 2016). Networks typically do not exceed a few centimeters in size, with many not exceeding 1 cm, and appear in growth position over, and blended into, micritic bands (fig. 5). Together, the micritic bands and sponge-spicule networks produced the familiar concave-down laminations that typically define protozoan stromatolites in general.
Beds 9 and 11 exhibit many factors consistent with an energetic, shallow, subtidal marine environment brimming with aquatic life. The bioclastic grainstones found in the spaces that separate individual forms in Bed 11, for example, are filled with trilobite hash and other invertebrate fossils such as brachiopod shells, mollusk shells, conodonts, radiolarians, sponge spicules, and even some echinoderm debris (fig. 4F).
Within the combined Cambrian/Ordovician carbonate succession both below and above the Hellnmaria Member of the Notch Peak Formation, there exist many other microbialite horizons, not including the eleven that I mapped while conducting my research (fig. 3).
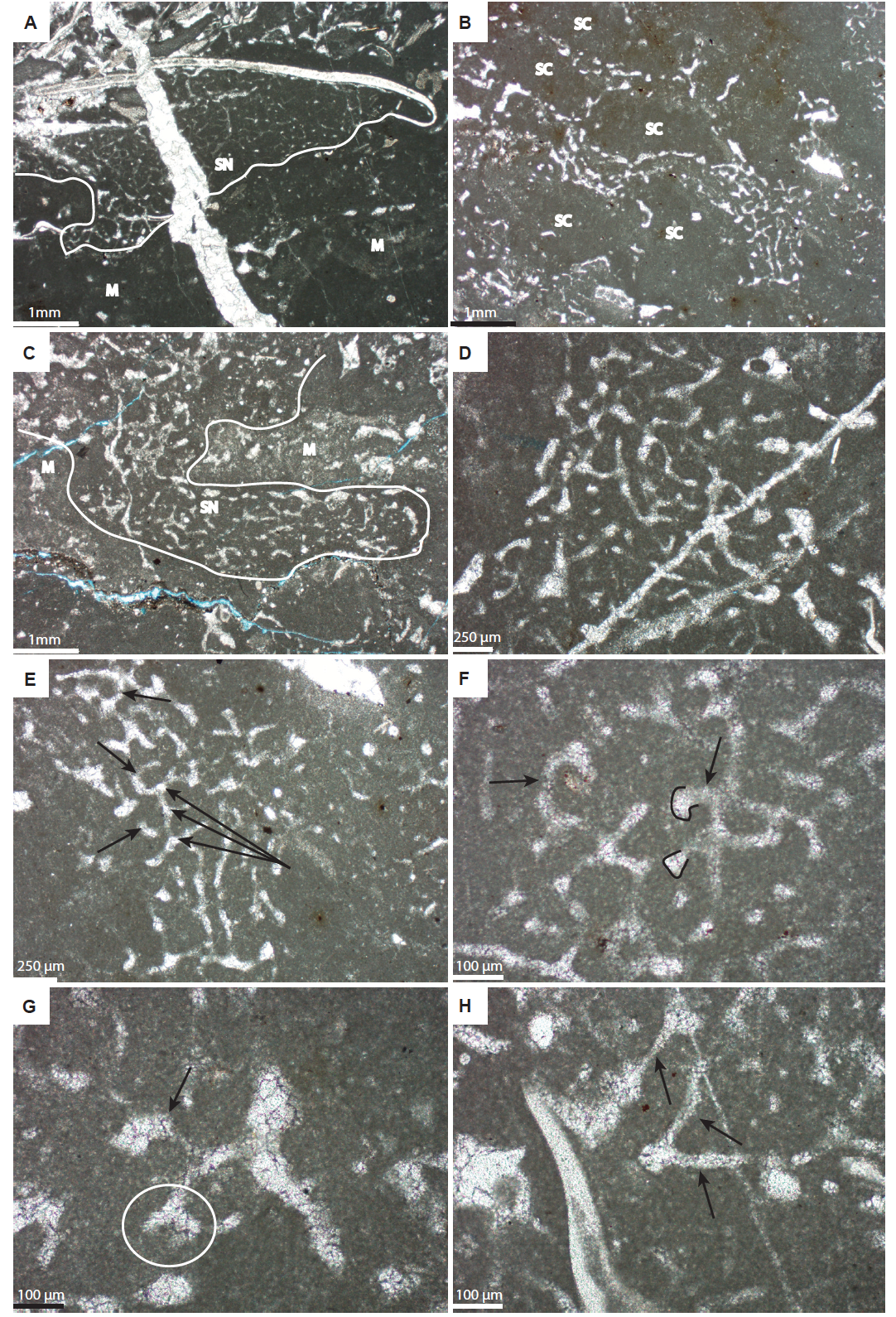
Fig. 5. Spicule networks from microbialites in bed 11. All images are in cross-section. Abbreviations: M = micrite; S = sponge; SC = sponge cavity. A. Sponge is attached to the underside of a trilobite carapace (under the “Shepherd’s Crook” which is diagnostic for trilobite carapaces). B. Spicule networks circumscribe possible sponge cavities (square enlarged in E). C and D. Two other sponge spicule networks. E. Desma-like spicules showing putative zygosis (long arrows) and curved, arcuate rays (short arrows). F. Partial network showing bulbous zygomes (outlined) and curved rays (arrows). G. Desma-like spicule showing possible gnarled zygome (circled) and curved ray (arrow). H. Spicules showing straight rays. Figure used with permission from SEPM, Figure modified and used with permission from SEPM, Coulson and Brand (2016).
Other Cambrian Microbialites in North America
Using the published literature, Lee, Sung, and Chough (2015) constructed a table of all known upper Cambrian microbialites from around the world. For North America, they described a total of 31 geographically distinct sites where upper Cambrian microbialites can be found. I went back over the papers referenced by Lee, Sung, and Chough (2015) for the purpose of mapping, and then differentiating apparently in situ microbialite assemblages from allochthonous ones (fig. 6). Of those 31 sites, 24 were interpreted by the authors to represent areas of in situ growth, conclusions with which I tentatively agree. In many of these locations, microbialites were found at multiple stratigraphic horizons. Some of the more extensive assemblages are summarized below.
In Newfoundland, an approximately 1,000 m thick succession of upper Cambrian through Middle Ordovician carbonate rocks record literally dozens of horizons of stromatolites and thrombolites. The stromatolites and the thrombolites are often enclosed within larger domed bioherms (having a domed shape) many meters in diameter. The upper Cambrian Berry Head Formation, for example, contains multiple different horizons of biohermal stromatolitic domal mounds composed of cabbage head (semi-columnar) to hemispheroid to digitate growth forms. Dozens of horizons of stromatolitic and thrombolitic buildups occur throughout the entire Cambrian-Ordovician succession and have been painstakingly detailed in multiple high resolution stratigraphic columns by Knight et al. (2008). All the domed buildups as well as individual columnar architectures were apparently right-side-up and in situ.
In Maryland, Demicco (1985) describes more than 20 microbialite-bearing horizons that themselves exist within a 3,700 m thick continuous sequence of Cambrian through Ordovician carbonates. Most of the microbialites were built by Renalcis-Girvanella calcimicrobial communities, have a thrombolitic clotted internal meso-structure, and often occur in 5 m thick biostromal beds (domes are joined together forming a more tabular reef). Although some of the microbialites in the cyclic facies (Frederick Limestone) are allochthonous, all of those found in the non-cyclic “lagoonal” facies (Conococheague Formation) are apparently in situ, and according to the stratigraphic column (Demicco 1985, 6), can have as many as ten separate horizons of meters thick microbialite assemblages per non-cyclic section (cyclic and non-cyclic sections are stacked one atop the other). Demicco (1985) correlated some of these beds over large distances of up to 50 km and interpreted many of these buildups as bioherms (so a domed top is assumed although I was unable to ascertain if the domes were right-side up). Since he does not describe inverted domes, and since he assumes in situ assemblages, I will tentatively concur with his interpretation.
In northeastern Tennessee, middle to upper Cambrian carbonates and shales have been interpreted as an intracratonic carbonate ramp by Srinivasan and Walker (1993). They describe multiple horizons (at least 3) of multi meters thick Renalcis-Girvanella bioherms within the middle Cambrian Maryville Limestone that can be correlated over about 60 km. These reefs are found within a single 40–120 m thick carbonate/shale cycle, of which there are at least three (see their sequence stratigraphic map on page 884). Srinivasan and Walker (1993) only discuss a single carbonate/shale cycle, so it seems likely that other reefs also exist within the other two carbonate/shale cycles. Again, no mention is made within the paper that any of these mounds were inverted.
In Texas, Johns, Dattilo, and Spincer (2007) describe multiple horizons (at least four) of multimeter thick microbialite sponge bearing reefs that span an 80 m section of the upper Cambrian Wilberns Formation. Two types of microbial reefs are present: a calcimicrobial-sponge assemblage that includes lithistid sponges and an eclectic assortment of the calcimicrobes, Girvanella, Epiphyton, and Renalcis, and a purely stromatolitic assemblage. The sponge microbial buildups are described as having “flat tops,” while a macromorphology is not provided for the purely stromatolitic forms. Sizes of these reefs vary from just 1 m high and 0.5–10 m in diameter, to as thick as 25 m for what they interpreted to be barrier reefs. There is no indication within the text that the stromatolitic columns were inverted.
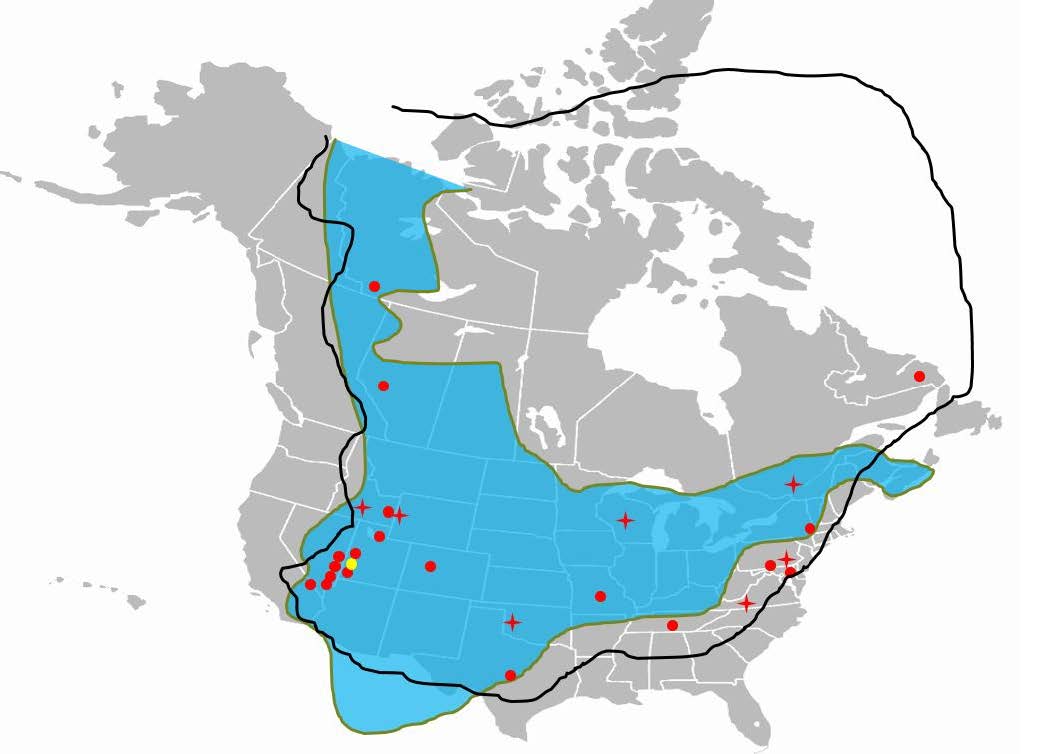
Fig. 6. Upper Cambrian microbialite distribution throughout modern-day North America. Red dots represent apparent in situ forms. Red stars represent either allochthonous distributions, or their in situ status could not be ascertained. Yellow dot represents the Notch Peak microbialites discussed in this paper. The blue U-shaped figure represents the interfingering of related Tapeats Sandstone equivalents. The black line represents the outline of Laurentia. Map public domain. Figure used with permission: Proceedings of the Eighth International Conference on Creationism, Coulson (2018).
In Colorado, upper Cambrian stromatolites that range in height from just a few centimeters to about 1 m have been described within the Dotsero Formation by Campbell (1976). This single horizon of columnar and hemispheroid stromatolites was correlated over an area greater than 1,000 km2. All forms are apparently right side up and in situ.
In correlated sections spanning eastern Nevada through western Utah, Shapiro and Awramik (2000) describe as many as ten horizons of meters thick microbialite assemblages. Stromatolites, thrombolites, and calcimicrobial derived (Renalcis) dendrolites of all different sizes and shapes are described in seven different 200–1,000 m thick sections. Some of these microbialite assemblages appear to correlate with microbialites from my own research site, and altogether constitute a large geographical distribution of reefs that grew on a Cambrian passive margin of staggering proportions.
In northern Utah, Saltzman et al. (2004) identified stromatolitic microbialite assemblages at six different horizons. Geographic scale and microbialite descriptors are not provided, except that some of these microbial buildups had thicknesses of about 3 m.
In the Canadian Rockies, Aitken (1967) describes a 450 km long, 350 m thick wedge of carbonate rocks that he interprets as a fossilized fore-reef, reef-crest/core, and back-reef. Much of the calcimicrobial (Epiphyton) reef core now exists as allochthonous fragments, but facies associated with the back-reef include multiple horizons (perhaps as many as eight) of apparently in situ stromatolitic microbialites.
These brief summaries are representative of all 24 locations in general. I have not personally seen any of these research sites but based on years of research from my own site, and on the information found within the papers, I see no reason to interpret them as allochthonous assemblages.
Importantly, almost all these locations fall upon a geographical arc that spans the coastline of ancient Laurentia (fig. 6). Starting in Newfoundland, this arc proceeds south to the New York/Maryland area before traveling through Tennessee, to Texas, and then to the regions around Utah, California, and Nevada, before continuing the trail northward through Idaho, Alberta, and into the Northwest Territories of Canada. Of these locations, almost all of them are stratigraphically located above Cambrian sandstones that themselves are thought representative of erosional processes related to the Great Unconformity (fig. 6).
Cambrian Microbialites in Korea
The 1,100 m thick Taebaek Group of eastern Korea is composed of 11 formations ranging in age from early Cambrian through Middle Ordovician. The 170 m thick Daegi Formation is the lowermost carbonate-bearing formation and sits atop the Jangsan/Myeonsan, and Myobong lower Cambrian cross-bedded sandstone formations, themselves unconformably sitting atop Precambrian crystalline basement rocks. Hong et al. (2012) describes 0.2–2.3 m thick siliceous sponge/calcimicrobe buildups that have lenticular to concave-down, as well as tabular geometries occurring at several levels through a 95 m thick section. The buildups are described as boundstones that are enclosed by well-bedded grey wackestone to skeletal packstone composed of sponge spicules, peloids, trilobites, and eocrinoid fragments.
Another reef-forming formation found within the Taebaek Group, but sitting stratigraphically higher than the Daegi Formation, is the 250 m thick Lower Ordovician Dumugol Formation which is composed of carbonate mudstones, packstones, and grainstones. Choh et al. (2013) describe 1.3–4.5 m wide and 0.7–1.9 m high, domed boundstone reefs, mainly comprised of stromatolites and thrombolites, with lithistid sponges to a lesser extent. Their lithographic column (Choh et al. 2013, 145) shows four separate horizons of large domed reefs intercalated with various types of carbonate sediments. The domed reefal bodies seem to be in a right-side-up position and are underlain by grainstones and packstones but overlain by mudstones.
Cambrian Microbialites in China
Large microbial/metazoan reefal complexes have been described by several workers from within the 1,800 m thick Cambrian-Ordovician succession of the North China Platform which covers 1,500,000 km2 (1,500 km east to west and 1,000 km north to south). At the base of the succession is the lower Cambrian sandstone Liquan Formation which sits unconformably upon Precambrian gneiss (fig. 7). The entire platform has been interpreted in terms of a vast epeiric inland ocean that existed from the early Cambrian through to the Middle Ordovician.
The combination lower to middle Cambrian Zhushadong and Mantou formations, which sit directly atop the Liquan Formation, contain 3–5 cm high and 5–10 cm wide, laterally-linked hemispherioid stromatolites (fig. 7). These smaller stromatolites are also found stacked on top of each other producing 10–80 cm high stromatolitic mounds. The stratigraphic columns of Lee and Chough (2011) describe at least six of these microbialite-bearing horizons spanning a few hundred vertical meters of carbonate sediments in both formations. Lateral extent of correlated microbial-bearing units is over 100 km.
The 180 m thick middle Cambrian Zhangxia Formation sits directly atop the Mantou Formation. Woo and Chough (2010) describe at least eight separate microbialite horizons intercalated between carbonate conglomerate sediments towards the top of the formation (fig. 7). Horizons are filled with 1.5–2.7 m high, club-shaped microbialites that widen towards the top and have a prominent radiating meso-structure. Microbialites are constructed of apparent colonies of the calcimicrobe Epiphyton that produced a solid, framestone “fan like” structure (fig. 1C). Individual microbialites coalesced with other microbialites laterally to produce multimeter thick, large scale biostromes that outcrop over many tens of kilometers. Based on the descriptions in the paper, all these microbialites are right side up and in apparent growth position. The reefal horizons are separated by carbonate conglomerates.
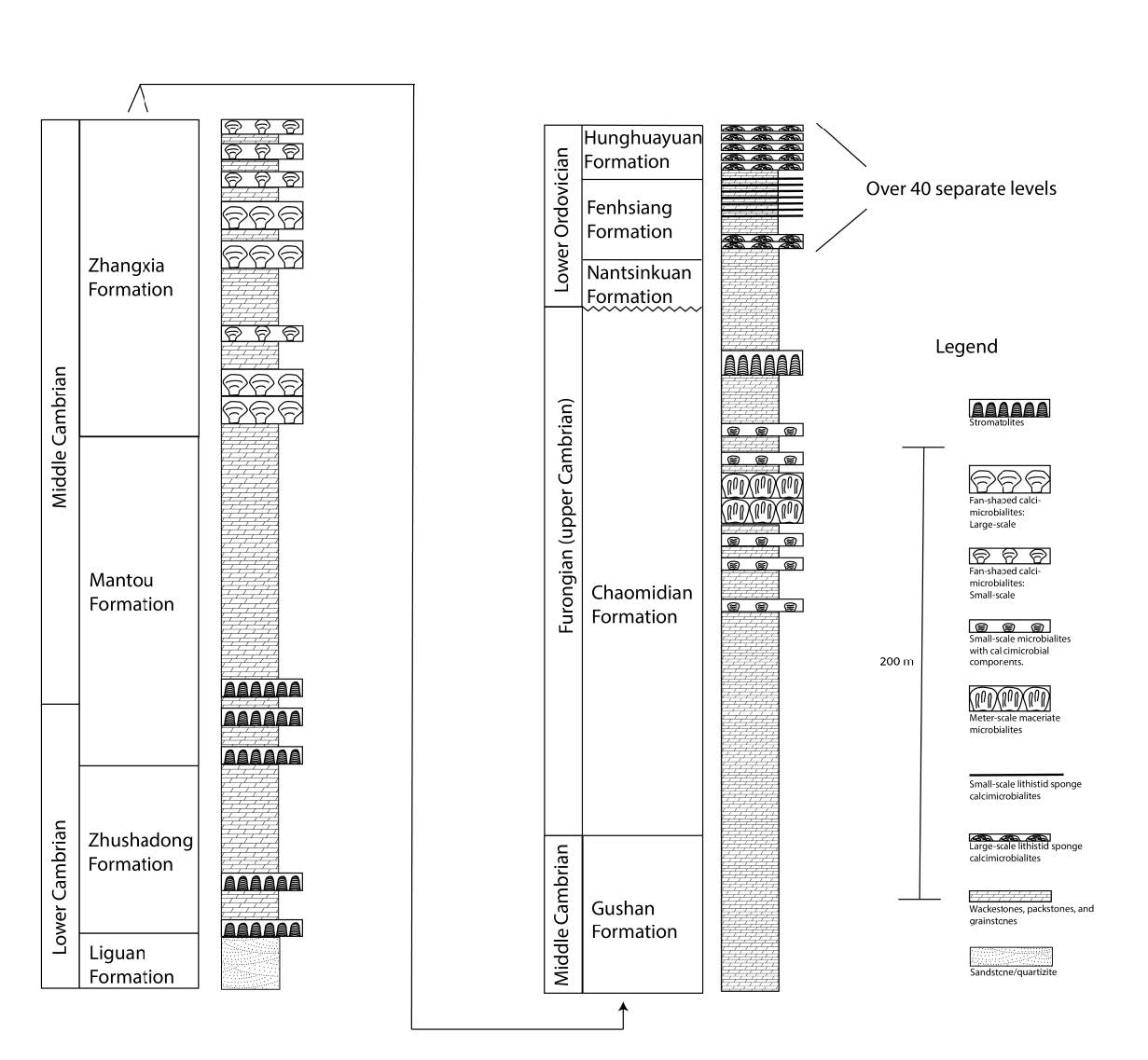
Fig. 7. Stratigraphic column of several Cambrian through Lower Ordovician rock sequences from China. Adapted from, Adachi, Liu, and Ezaki (2013); Chen, Lee, and Woo (2014); Lee, Chen, and Chough (2010); Li, et al. (2015); Lee and Chough (2011); and Woo and Chough (2010). Box size encapsulating the microbialites shows only a relative thickness to other microbialite beds. The position of the microbialites within each formation is only approximate but powerfully demonstrates the existence of dozens of apparently in situ, meters-thick microbialite beds intercalated between great thicknesses of carbonate sediments.
Sitting stratigraphically above the Zhangxia Formation is the 190–260 m thick upper Cambrian Chaomidian Formation. Chen, Lee, and Woo (2014) describes three different kinds of microbialite reefs within this interval (fig. 7):
- Small-scale microbial reefs constructed by the calcimicrobes Girvanella and Renalcis that form small domes a few centimeters to a few decimeters in height and occur sporadically at multiple horizons throughout the formation. Small digitate stromatolites also make up the mesoscale fabric of some of these microbialites.
- Meter scale maceriate microbialites are composed of centimeter-scale “maze-like” structures (hence the name maceriate). Each maceria (singular) follows a branching, column-like shape in longitudinal section and a maze-like shape in transverse section. These reefs have a typical dome-shaped architecture and are often coalesced with other domes forming compound biostromal complexes. A single 10–20 m thick horizon of maceriate microbialites was correlated over 100 km by Lee, Chen, and Chough (2010) and has an estimated overall areal distribution of about 6,000 km2.
- Purely stromatolitic reefs can be found towards the top of the Chaomidian Formation. Stromatolites exist as columns that are commonly 30–100 cm in height and about 10–50 cm in diameter. One of these beds was traced for several tens of kilometers.
Chen, Lee, and Woo (2014) furnishes a thorough stratigraphic column that has six different sections extending over about 500 km. One of these sections has five different reefal horizons. When domed, the buildups are always right side up, as are all the stromatolites that form the stromatolite-specific horizon at the top of the Chaomidian Formation. All the reefal horizons are separated by skeletal packstones and wackestones.
Moving further south (about 400–600 km south of the succession on the North China Platform), Adachi. Liu, and Ezaki (2013) describe sponge microbial calcimicrobial, sponge bryozoan, and bryozoan pelmatozoan framestone reefs that span five lithostratigraphic Lower Ordovician units on the South China paleoplate (fig. 7). Their stratigraphic column (Adachi, Liu, and Ezaki 2013, 454) shows the existence of reefs on at least 40 different levels that span 75 m. Most of these reefs are quite small, perhaps just a few decimeter in height and/or thickness, but more than ten horizons have large domed reefs that are 2–6 m in diameter and about 1–2.5 m in height (Adachi, Liu, and Ezaki 2013; Li, et al. 2015). Adachi, Liu, and Ezaki (2013) compared these reefs to similar sponge microbial bryozoan reefs at two different locations on the Yangtze Platform. All three reefal sections were separated by hundreds of kilometers but retained similar paleobiological characteristics. They concluded that all three locations show a transition from microbe-dominated reefs at the bottom of each section, to sponge stromatolite reefs to sponge-calcimicrobial reefs to sponge bryozoan reefs at the top of each section. As with the North China Platform, the Yangtze Platform has been interpreted in terms of an extensive epeiric sea. Also, along with the reefs discussed above, all domed structures appear to be right-side-up and covered over with once unconsolidated bioclastic/skeletal grainstones and packstones.
Cambrian Microbialites Elsewhere
Although North America and China boast some of the best upper Cambrian microbialites in the world, they are not the only places where abundant Cambrian microbialites can be found. Plentiful microbialite beds can be found in Russia, Australia, Iran, Argentina, Kyrgyzstan, and even Antarctica (Hong et al. 2012; Lee, Sung, and Chough 2015).
Discussion
Until recently, most creationists assumed that Paleozoic microbialites, in comparison to Precambrian assemblages, are rare or occur to a much lesser extent (Clarey 2018; Purdom and Snelling 2013; Snelling 2009). Yet Cambrian and Ordovician microbialites are quite common, and according to Peters, Husson, and Wilcots (2017), “transiently achieved Proterozoic-like prevalence during the Paleozoic” (emphasis mine). Clearly, this perception within the creationist community needs to change. Paleozoic microbialites are abundant.
These data, however, conflict with current creationist models of earth history, especially as it relates to the onset of Noah’s Flood. Most creationists place the pre-Flood/Flood boundary at or near the Precambrian/Cambrian boundary (Austin and Wise 1994; Dickens 2017; Dickens and Snelling 2008; Snelling 2009). This means that Cambrian sediments represent Flood rocks. Yet how can so many microbialites grow on dozens of stratigraphic horizons to such great thicknesses over large geographical areas all over the world in just one year? Since many creationists incorporate the entire Paleozoic and Mesozoic into the Flood year, then Cambrian rocks would only represent a small portion of that year—perhaps just a few months. The ramifications for this position are obvious and striking.
Criteria for Establishing an in situ Origin for Stromatolites
Did these microbialites really grow in place or were they moved to these locations during the Flood? Another way of saying this is, are these microbialites autochthonous (in situ) or allochthonous (transported)? These are very important questions, and as such, require specific criteria that will furnish conclusive answers. A list of nine criteria demonstrating an in situ origin for the Utah stromatolites will now be discussed.

Fig. 8. Multiple stromatolites that clearly show an organic attachment to the substrate. A. Stromatolite and substrate removed from Bed 11. B. The same stromatolite from A in cross-section capturing the internal stromatolitic fabric, the interspace material, and the stromatolite’s attachment to the substrate. Varying stages of growth can be seen within the stromatolite. C. Another stromatolite that took advantage of the protruding substrate is seen with concave-down growth laminations radiating away from the substrate. The stromatolite is clearly draped over this protrusion and then fans out into a widening-upward columnar stromatolite. D. This stromatolite is clearly attached to the substrate. Although meso-structure laminations cannot be seen, the distinct widening-upward club-like macro-structure is striking. E. Another stromatolite that has taken advantage of the elevated substrate. Notice that the concave-down laminations have grown over the protruding substrate as with the stromatolite in C. F. This is an interesting photo in that it shows stratiform (growing more laterally over the substrate than upward like the columnar forms) stromatolites growing not only from the substrate, but from one another. G. Although not as conclusive as the latter stromatolites, these three stromatolites can still be seen growing from the substrate. With thanks to Ronny Nalin for photos in D and G.
The most straightforward data supporting an in situ growth model for the Utah stromatolites is the attachment of the structure to the substrate. This criterion doesn’t require a great deal of explanation; if the stromatolites are in situ, then evidence for that in situ growth should be evident in the form of an organic connection with the substrate. If this criterion can be satisfactorily established, then this in most instances should automatically preclude an allochthonous origin (see below for transport within megaclasts). Fig. 8 shows multiple stromatolites that persuasively fulfill this criterion.
One of the most important examples is represented in fig. 8A. This is because this entire stromatolite, including the substrate consisting of a large 100–150 lb block, was removed from the Utah site for sectioning allowing me to conduct a thorough investigation. The stromatolite was cut along the black vertical line which was indicative of the stromatolite’s and the substrate’s center. The arrows represent the interspace material that was still in place when we found this particular specimen. The stromatolites that closely neighbored this one can be seen in fig. 9D. All the stromatolites at this particular location are weathering out of the limestone layer providing a unique 3D perspective that captures something of their original setting. Fig. 8B shows the same stromatolite in cross-section exposing the internal stromatolitic fabric, the interspace material, and the stromatolite’s attachment to the substrate. Notice that the stromatolite is attached to an elevated section of the micritic substrate (arrow) which was most likely a lithified hardground.2 The basal section of the stromatolite lies between the solid lines, which itself is buried in the very coarse interspace grainstone. Varying stages of growth can be seen within the stromatolite. The first growth stage has been traced in as a narrow concavity. Radiating out around this initial growth stage are two later stages that clearly depict concave-down laminations typical of stromatolites (also traced in). The inset image in fig. 8B shows the same area without tracings. A fourth line has been drawn in the lower-middle part of the stromatolite for clarity, showing that these concave-down laminations contributed to the overall widening-upward shape.
In fig. 8C, another stromatolite that took advantage of the protruding substrate is seen with concave down growth laminations radiating away from the substrate. The stromatolite is clearly draped over this protrusion and then fans out into a widening-upward columnar stromatolite. Another smaller stromatolite is outlined to the right. This one has grown from the flat surface and shows that these stromatolites, although preferring elevated surfaces, can grow over a hard, flat substrate. These two stromatolites actually connect farther up the wall and are seen as almost flat laminations towards the top of the image.
The stromatolite in fig. 8D, as with the latter two forms, is clearly attached to the substrate. Although meso-structure laminations cannot be seen in this image, the distinct widening-upward club-like macro-structure is striking. This stromatolite grew on a very coarse, and sometimes cross-bedded ooid grainstone.
Another stromatolite that has taken advantage of the elevated substrate can be seen in fig. 8E. Notice that the concave-down laminations have grown over the protruding surface as with the stromatolite in C.
Fig. 8F is an interesting photo in that it shows stratiform (growing more laterally over the substrate than upward like the columnar forms) stromatolites growing not only from the substrate, but from one another! Stromatolite 1 has taken advantage of the cone-shaped elevation and grows laterally out in both directions. At some point during growth, strong currents or a storm deposited some wackestone infill. Then at another point in time, the second stromatolite has grown both over stromatolite 1 and the wackestone infill. The stromatolites in fig. 8G are not as convincing as the others, but the growth of three stromatolites from the surface is still fairly conclusive. Interestingly, the interspaces between these three forms are quite wide. Most interspaces have a width of about 10 cm, but this is because they are immature forms (see also below).
-
The second criterion that establishes an in situ origin for the Utah forms is the consistent upright orientation of these stromatolites. While conducting my research at the Utah location, I had the privilege of seeing literally hundreds of these structures. In every instance, through all eleven horizons and over about 20–40 km2, all of these stromatolites, except one, were upright. This can be fully appreciated in fig. 9G. This aerial photo was taken by a drone and shows hundreds of stromatolites from a top-down view. Most of these stromatolites are around 50–70 cm in diameter and are evenly spaced. This spacing (discussed further below) is a consistent feature for all of the stromatolites in the Utah location at all 11 horizons over the entire research area. Fig. 9D provides a stunning 3D perspective of what these stromatolites might have looked like as they grew from the ocean floor. The upright nature of these stromatolites is used as a criterion because I fail to see how these structures could have been deposited in an upright orientation given a very energetic depositional environment (the Flood). The laminated portions of these stromatolites are micritic, although thrombolitic wackestone pockets can be found towards the middle/top of some of them (fig. 4B). As can be seen from fig. 8B, this micritic texture is quite dense; there are very few vugs or spaces in the forms from Bed 11,3 although fenestral domains do exist and are interpreted as deteriorated sponges (Coulson and Brand 2016). Most of the Utah stromatolites are also taller than they are round, furnishing them with a “barrel-like” shape.
All introductory geology textbooks clearly state that solid objects with long-axes are always transported with the long-axis parallel to the ground. In other words, solid “barrel-like” objects are not transported in an upright orientation. If these stromatolites represent a transported assemblage, then how did they manage to either stay upright or “get” upright once they arrived in Utah? Thankfully, I did get to see and photograph a single allochthones stromatolite. Fig. 9C shows a stromatolite caught in what appears to be a wackestone mudflow with the stromatolite’s wider head located below the narrower pedestal. According to Wise and Snelling (2005), microbialites that widen-upward are best interpreted as in situ if the pedestal is found below the head. This makes sense since gravity would tend to position the heavier, wider head towards the bottom if in fact the form was transported. This example fits that criterion. Wackestone also surrounds the entire structure bearing testimony to its allochthonous origin. If transported, we should expect to see other stromatolites deposited in a manner similar to this.
Some colleagues have suggested that these stromatolites might have individually “floated” to this location. But most of these structures are almost solid micrite having a density of about 2,700 kg/m3. Since water has a density of about 1000 kg/m3, such a “floating” hypothesis is difficult to imagine.
The third criterion is closely allied to the second. Since modern stromatolites grow one lamination at a time in a concave-down orientation (Coulson, Brand, and Chadwick 2016), we have good reason to suppose that the same was true for ancient forms. As far as I know, there is no such thing as a columnar stromatolite that has concave-up laminations. If these stromatolites are indeed in situ, one would expect to find consistent concave-down laminations across the research area. If these stromatolites were transported individually, we should expect to see an inconsistency whereby half of the stromatolites were deposited upside-down. Even if the stromatolites had some kind of a preference to “float” at a particular orientation (perhaps because one end was less dense than the other), then some of the stromatolites should still be expected to be deposited upside-down. Yet the stromatolites are consistently, 100% of the time, orientated in a concave-down fashion. This orientation is thus strong evidence for in situ growth.
-
The fourth criterion validating in situ growth is the presence of very narrow interspaces that completely surround the structures. In fig. 10 A–D, various kinds of stromatolites are pictured from a top-down perspective. Notice that the macro-shape and even the internal fabric of each suite is different in each image. These different internal fabrics and external shapes most likely resulted from differing microbial communities. Remarkably, all the interspace areas, no matter the macro-shape, are very narrow—on the order of just a few cm to perhaps as many as 20 cm wide. How likely is it that transported stromatolites could be randomly deposited so as to maintain consistent interspaces of just a few centimeters? This observation is constant for thousands of stromatolites covering about 20–40 km2. Transported stromatolites would certainly not maintain such consistent spacing and should be found in closely packed groups showing evidence of collision.4 There is actually a reasonable biological explanation for this incredibly uniform distancing. Petroff et al. (2010) observed the same phenomena in conical microbial communities in Yellowstone National Park. They used these stromatolites as a proxy to build a distancing model of growth based on microbial competition for nutrients. They say: “We note that all modern conical stromatolites and many that formed in the last 2.8 billion years display a characteristic centimeter-scale spacing between neighboring structures. To understand this prominent—but hitherto uninterpreted—organization, we consider the role of diffusion in mediating competition between stromatolites.” The three boxes in fig. 11 provide a synopsis of their model. Essentially, given an environment that is conducive to abundant microbial growth, stromatolites, although spaced randomly at first, will tend to grow laterally in every direction (hence the round shape). This kind of growth and shape ensures the greatest possibility of obtaining the most nutrients. But as stromatolites grow closer to each other, the nutrients begin to dry up and a zone of competition is established. Although I’ve simplified their model, the overall result is a tightly packed field of stromatolites that seldom touch.
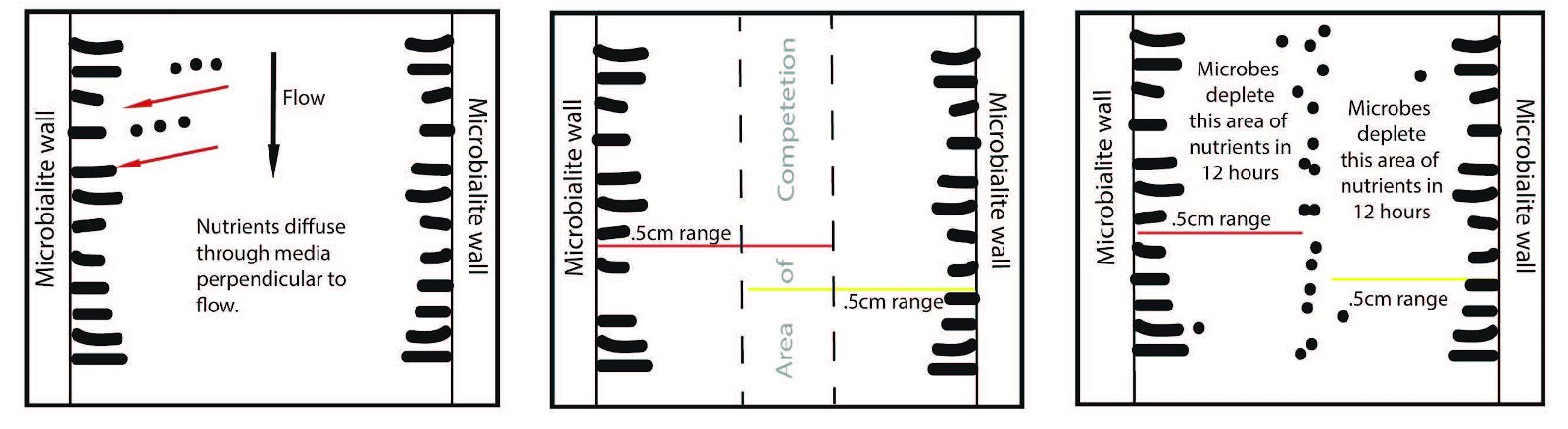
Fig. 11. These three boxes are adapted from Petroff et al. (2010) and represent a very simple version of their nutrient diffusion and competition model.
Caution must be applied when directly applying these data to the Utah forms since the microbialites growing in Yellowstone National Park are freshwater examples forming in the absence of sedimentation and are many orders of magnitude smaller, but the researchers do try to apply their model to the fossil record and propose some modifications that may help explain the much larger interspace gaps found in fossilized forms (Petroff et al. 2010, 9960). They hypothesized that competition for nutrients might continue to influence interspace gaps, but on a larger scale, given the role of molecular diffusion in concert with moving water and advection.
This case study provides convincing evidence supporting in situ growth for the Utah stromatolites. I think that this interpretation is most strongly realized in the shapes of the more dendrolitic thrombolites in fig. 10B. According to Petroff et al., when living communities of microbes compete for space, their macro-shape becomes somewhat hexagonal:
“In still water, each structure competes with neighbors on all sides. A field of stromatolites then reaches a maximum density while preventing direct competition by growing into a hexagonally symmetric pattern. . . . Indeed, we often observe that nascent clumps form into a roughly hexagonal arrangement. . . . Because these aggregates grow on smooth surfaces such as glass beakers, the regular spacing must be intrinsic to the growth of these biofilms, rather than inherited from the topography of the underlying surface. A similar hexagonally symmetric arrangement can also be observed growing on the surface of a mat that had been deformed by a gas bubble.”
The hexagonal macro-shape is clearly evident in fig. 10B. Given that these stromatolites have the narrowest interspaces, as little as 1–2 cm between forms, it is entirely possible that these stromatolites took on this hexagonal shape in conformity to the research outlined above. See also Tóth (1948).
An allochthonous model not only has the uphill battle of explaining how transported stromatolites somehow managed to consistently maintain narrow interspaces, but it must do so while disregarding a viable growth alternative.
The fifth criterion establishing an in situ interpretation for the Utah stromatolites is associated with the elongate forms. Although not as common as the roundish forms, elongate stromatolites can be found throughout the research area. These elongate forms are typically 1–2 m long and a few to as many as 4 m tall, but only average widths of about 20 cm (fig. 10D). Some of these elongate forms are up to 5 m long!(fig. 9A). If these elongated stromatolites had been transported, then how did they remain stable while sliding along on a 20 cm wide base? Remember, they are several meters long and tall. The problem is compounded when one considers the consistent narrow interspaces that border all of the stromatolites (fig. 9A and B). How did these stromatolites not only remain upright, but how did they come to “park” themselves nicely next to their neighbor while maintaining consistent distances of just a few cm? The most parsimonious and simple explanation is that these elongate forms grew in place. The narrow interspaces are also consistent with the growth model discussed by Petroff et al. (2010). Although still unclear, it seems reasonable to conclude that the stromatolites took on a preferential elongated shape in the presence of a low to moderate flow regime, something I discuss in my original research (Coulson, Brand, and Chadwick 2016).
The sixth criterion establishing an in situ interpretation for the Utah stromatolites is the interlocking nature of some of the forms. Fig. 10C shows unique stromatolites that took on some rather bizarre shapes. I am not exactly sure why this occurred, but what becomes immediately obvious is the interlocking nature of these shapes. I cannot conceive of any allochthones model that would pick up uniquely shaped stromatolites, transport them to another location, and then proceed to interlock them in the same puzzle-like form they had in their original setting! Only an in situ growth model can account for such an arrangement.
-
The seventh criterion validating an in situ growth model is the presence of an inter-connecting suite of stromatolites over several kilometers. In fig. 2, to the right of Bed 9 in the stratigraphic column, I have sketched an upward series of growing stromatolites. This sequence changes shape from round on the bottom, to elongate in the middle, to round again at the top, and is consistent across all seven outcrop locations. Essentially, the round stromatolites at the bottom of the series entered a bidirectional flow regime that, in connection with the nutrient diffusion model above, caused the morphology to change from round to elongate. The top of the series represents a reversing of this trend as the bed exited this flow regime. This vertical series can be clearly seen at all seven locations which means that the entire suite of forms is laterally connected (see Coulson, Brand, and Chadwick 2016, fig. 3). In other words, the round forms appear on the bottom at all seven locations, the elongate forms appear above the round forms at all seven locations, and the round forms again appear over the elongate forms at all seven locations. The series sketched in fig. 2 shows that intermediate morphs also occur between the round and the elongate forms and between the elongate and round forms on top. This occurs across 8 km. If these stromatolites were transported as individual structures, then how did they manage to stack themselves in this pattern over the entire research area? Only two alternatives exist for this particularly thick reef (averaging 10 m). Either it grew in place or the entire structure was transported as a single, 8 km long carbonate clast. Since no evidence for the latter interpretation is present at the research site, the most parsimonious solution is that this entire reef grew in place.
-
The eighth criterion establishing an in situ origin for the Utah stromatolites is the presence of narrow bridges that link some of the structures. Figs. 9F and 10D respectively show round and elongate stromatolites that are connected by narrow microbial bridges (white arrows). Although microbial competition tended to restrain microbial amalgamation, this doesn’t mean it did not occur. These microbial bridges are actually quite common and can best be explained in an in situ growth model, but cannot be explained in one that requires transport, especially transport during an energetic depositional environment (the Flood).
The ninth criterion establishing an in situ origin for the Utah stromatolites is the petrographic correspondence that exists between the stromatolites and the interspaces in Bed 9, and between the interspaces and the substrate in Bed 11. Bed 9 records a changing microbialite morphology from round at the bottom of the bed to elongate and then back to round again at the top of the bed (fig. 2). This changing morphology resulted from the introduction of a shallower-water bidirectional flow regime. The round shapes at the top of the bed resulted from the bed subsiding back into deeper water and exiting this flow regime. The petrographic data support this interpretation. The interspace sediments for the round microbialites at the base of the bed are characterized by the abundant presence of micrite (fig. 12A). Trilobite bioclasts are randomly located throughout the matrix, constitute ≈ 25% of the allochems and are ≈ 250 µm to ≈ 3 mm in length. Small packstone pockets of medium to large-sized intraclasts up to ≈ 1.5 mm in size are also present. All of the allochems are angular and poorly sorted. This material is best described as a bioclastic wackestone with some intraclastal packstone domains, and is interpreted in terms of a deeper water, low energy environment.
The interspace sediments for the elongated forms, however, are composed of moderate to well-sorted, well-rounded, sub-spherical, micritized intraclasts ≈ 250 µm to 0.5 mm in diameter, and make up 80% of the allochems (fig. 12B). Trilobite fragments constitute less than 20% of the allochems, and are typically less than 0.5 mm in length. Some intraclastic packstone domains are randomly scattered throughout the matrix. These interspace sediments are best described as intraclastal grainstones with some intraclastic packstone domains, and are interpreted in terms of a moderately energetic environment—the elongate morphology resulting from a bidirectional flow regime.
The interspace sediments for the round microbialites at the top of the bed are composed of ≈ 20% trilobite bioclasts, ≈ 250 µm to several millimeters in length (fig. 12C). Intraclasts ≈ 0.5 to ≈ 2 mm in diameter appear in isolated packstone pockets and constitute ≈ 30% of the allochems. All of the allochems are angular and poorly sorted, as was the case with round forms at the base of the bed. These sediments are best described as bioclastic wackestones, and are interpreted as a deeper water, low energy environment consistent with subsidence.
The petrographic data associated with Bed 11 is also pertinent. The stromatolites in this bed grew in very shallow water, and differ from the other 10 beds by the presence of well-washed grainstones. Unlike Bed 9, the interspace material in Bed 11 is a well-washed bioclastic grainstone composed of trilobite hash (fig. 4F). Micrite is rare. The entire bed is also sitting on a well-washed ooid grainstone (fig. 12D) that regularly exhibits herringbone cross-beds (fig. 9E).
The petrographic data from both beds fit an in situ model of stromatolite growth but do not accord with an allochthones model at all. The micritic, poorly sorted wackestones found in the interspaces of the round stromatolites from the bottom and the top of Bed 9 fit with a low energy environment, while the moderate to well-sorted, well-rounded, subspherical, micritized intraclasts of the elongate layer fit better with a more energetic environment—a good fit with the elongate form itself. It seems to stretch the imagination to conceive of allochthonous processes producing these correlatable petrographic relationships.
The same is true for Bed 11 and is perhaps even more striking. Why would the stromatolites be sitting on an ooid grainstone substrate yet have completely different interspace allochems (bioclastic hash) if the entire suite of sediments and stromatolites were moved in an energetic Flood? Surely things would get mixed up? It makes more sense to interpret the bioclastic hash found in the interspaces of the stromatolites in terms of a shallow-marine environment thriving with animal life, and the ooid grainstone as the substrate upon which they grew. The herring-bone-cross-beds (fig. 9E) are also at odds with an allochthonous model, especially one producing a highly energetic unidirectional flow over vast distances. These low relief, herring-bone-cross-beds are better interpreted in terms of a shallow-water, intertidal environment that experienced the ebb and flow of bi-directionality. The direction of flow can even be determined from the cross-bedding dips (Coulson, Brand, and Chadwick 2016).

Fig. 9. Additional stromatolite information. A. Very long—sometimes 5 m—elongate stromatolites. These elongate forms are separated by consistent, narrow interspaces, are typically several meters high, but only have decimeter-scaled widths. B. A cross-sectional view of an elongate stromatolite. Notice that the narrow spacing is consistent from the top to the bottom. C. An allochthones stromatolite, perhaps caught in a mudflow, or ripped up in a storm and deposited in a muddy matrix. Notice that the wider head is below the narrower pedestal. D. Stromatolites from Bed 11 are weathering out of the bed and can be viewed in 3D and in growth position. E. Herring-bone cross-bedded ooid grainstone bears testimony to the bi-directional, and most likely tidal, flow regime. Bed 11 stromatolites are growing on this substrate. F. Two round stromatolites are connected by a microbial bridge. G. An arial photo taken by a drone showing hundreds of stromatolites in growth position. Although difficult to see, the whole image is filled with stromatolites. White arrows point to specific stromatolites. For scale, the white arrows are about 2 m long.

Fig. 10. Narrow interspaces are consistent across each stromatolite bed, and at every horizon within the Hellnmaria Member. These plan-view photos were taken at four different horizons. Differing macro and mesostructures most likely resulted from differing microbial communities in conjunction with varying water depths and sub-aqueous flow regimes. A. Bed 11 (fig. 2) stromatolites are typically round to slightly oblong and are stromatolitic. B. Bed 10 (fig. 2) microbialites are thrombolites with a dendritic branching meso-structure. These forms were tightly packed together having extremely narrow interspaces—just 1–2 cm. Notice the hexagonal-like macro-shapes. C. Bizarrely-shaped microbialites (mesostructure could not be determined) that “interlock” like a jigsaw puzzle. D. Elongate stromatolites from Bed 9 (fig. 2). Arrow is pointing to a microbial bridge connecting two structures.
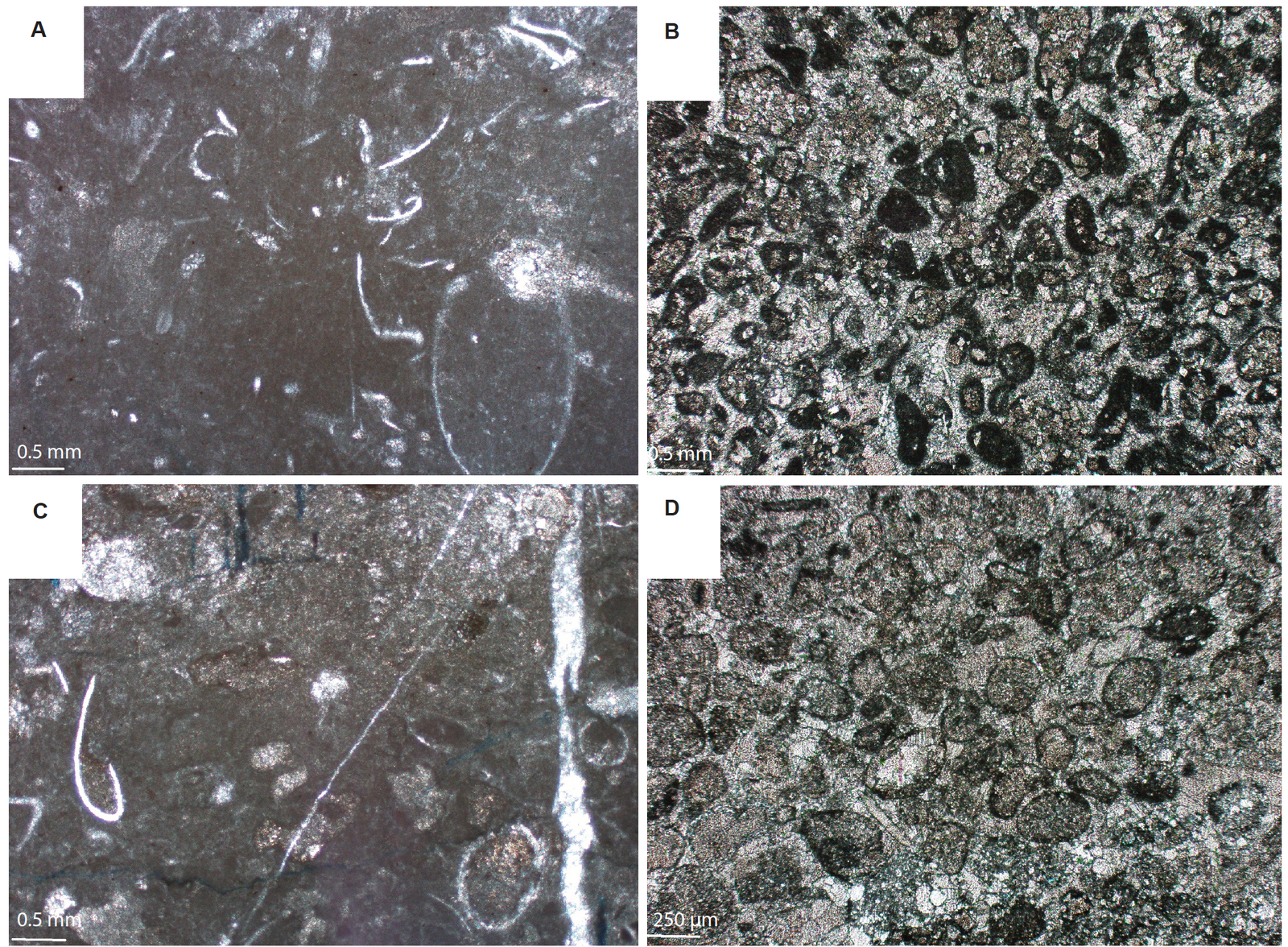
Fig. 12. Petrographic data. A. Bioclastic wackestone from the round stromatolite interspaces at the base of Bed 9. Micrite is abundant (fig. 2). B. Moderately to well rounded, moderately sorted intraclastic packstone with wackestone domains from the interspaces of the elongate forms in the middle of Bed 9 (fig. 2). C. Bioclastic wackestone from the round stromatolite interspaces at the top of Bed 9 (fig. 2). D. Well-rounded, well-sorted ooid grainstone from the substrate of Bed 11. Adapted from Coulson, Brand, and Chadwick 2016.
Although not useful as an in situ growth criterion, the presence of encrusting sponges in some stromatolites does bear testimony to growth in general. This is important for those who think that these stromatolites were not biologic structures. Sponges are recognized as spicule networks that typically do not exceed a few cm in size, with many not exceeding 1 cm (fig. 5). Spicule networks display various morphologies, although a laminated form is the most common, and occupy about 50% of the surface area when seen in cross-section. The millimeter thick micritic lenses crudely intercalated between the spicule networks tend to be homogenous, blotchy light to dark gray in color, and locally host abundant bioclasts. Very fine wavy to horizontal micritic laminae, small convex micritic mound-like, and micritic fan-like structures are also randomly distributed throughout these micritic lenses which for the most part are structureless. Calcimicrobes, typically identified on the basis of diagnostic genera such as Epiphyton, Girvanella, and Renalcis, are absent from thin sections.
I proposed a growth model whereby microbial biofilms first colonized and stabilized the underlying substrate. As a result of continued microbial trapping and binding of lime mud and/or precipitation of micrite, the fabric acquired a stromatolitic texture. This initial rigid microbialite with pre-existing concave-down laminations served as a suitable substrate for early sponge attachment (Kruse and Reitner 2014). From that point onwards, microbial and sponge communities reciprocally encrusted and immured each other, resulting in the construction of columnar “stromatolites” that retained a concave-down laminated meso-structure.
Antediluvian Environment or Flood deposits?
Having established an in situ interpretation for the stromatolites, it will now be important to establish a model of deposition that is consistent with the antediluvian world. Creationists Whitmore and Garner (2008) compiled a list of criteria that creationists could use to differentiate Flood deposits from pre- and post-Flood deposits (table 1). After applying their criteria to the Notch Peak Formation, my initial results were rather inconclusive. Boiling it down to the simplest common denominator, this is most likely due to the presence of two conflicting criterions—the presence of marine deposits over the continents and the presence of reefs. In Whitmore and Garner’s paper, marine sediments over the continents are given a high priority in favor of Flood deposits. But that criterion may not be valid. It is true, a catastrophic Flood should deposit marine sediments upon the craton, but so could processes working over a pre-Flood epeiric inland sea. We have examples that we can point to today to justify that possibility. Whitmore and Garner even discuss that possibility on page 428: “Some widespread deposits may also have formed, however, during the regression of waters from the continents on Day Three of Creation Week and in the pre-Flood epeiric oceans” (emphasis mine). Since the existence of epeiric inland seas during the antediluvian period remains a biblical and scientific possibility, then I do not think we can use this criterion as prima facia evidence supporting a global Flood.
| X | Marine deposits on the continents. r = 1 |
| ? | Deposits of unparalleled extent. r = 1 |
| ? | Global and regional unconformities. r = 1 |
| X | Transgressive sequences. r = 2 |
| Delta deposits. r = 3 | |
| ? | Mass kill deposits. r = 2 |
| Coal deposits. r=2 | |
| X | Last appearances of extinct marine species. r = 2 |
| X | Seawater temperature. r=2 |
| High sea level. r = 2 | |
| Geological energy. r = 2 | |
| Tectonic activity. r = 2 | |
| Volcanic activity and deposits. r = 3 | |
| X | Original horizontality preserved. r = 3 |
| Local sedimentary units. r = 3 | |
| X | Bioturbation. r = 3 First appearance of e |
| X | First appearance of extant species. r = 2 |
| Lacustrine deposits. r = 2 | |
| Fluvial deposits. r=3 | |
| X | Regressive sequences. r=2 |
| Widespread true glacial deposits. r = 1 | |
| X | Evolutionary species diversity. r = 1 |
| X | Large in situ reef structures. r = 2 |
| Terrestrial vertebrate trackways. r = 2 | |
| True desiccation cracks. r = 3 | |
| True evaporite deposits. r = 3 | |
| Aeolian deposits. r = 2 | |
| True paleosols. r = 3 | |
Once this criterion is removed, however, the rest of the criteria do seem to support either a pre- or post-Flood interpretation. I put a question mark in for “deposits of unparalleled extent” and “global and regional unconformities” because I only assessed the Notch Peak Formation (table 1). Although this Formation does not possess regional deposits and unconformities, Cambrian sandstones that lie far below the Notch Peak Formation actually do. Again, however, if we allow for antediluvian inland epeiric seas, then “deposits of unparalleled extent” must also be removed as a primary criterion. Since major “global and regional unconformities” would be expected during Creation Week, this criterion must also be interpreted very carefully. In the model proposed in this paper, there are no scientific or biblical reasons to demand a “tick” in these first three bins. Looking at the bottom of the table,5 however, and the presence of reefs, evolutionary species diversity (trilobites), and bioturbation seem to require a pre- or post-Flood interpretation. You simply cannot have extremely large reefs, for example, growing in just a few months. “Mass kill deposits” also received a question mark because of how the presence of bioclastic hash in the stromatolite interspaces could be interpreted. I contend that this bioclastic hash is best interpreted as trilobite exuviae and abandoned shells typical of any thriving shallow-water environment.
That a diverse marine environment once flourished throughout the western and central United States is also supported biostratigraphically. Fig. 13 compares two suites of trilobite zones, one suite from the upper Cambrian Notch Peak Formation in Utah, and one suite from the Llano Uplift in Texas. Both suites contain eight trilobite zones that not only correlate but correlate in their relative positions within the sediments. Notice that these eight zones do not appear out of order. Instead, each zone corresponds with the other in both trilobite type and relative position. Taylor et al. (2012) have compiled a list of several other western and central North American locations including Northern Canada, Nevada, California, Colorado, New Mexico, and Oklahoma, that exhibit a very similar biostratigraphic sequence.
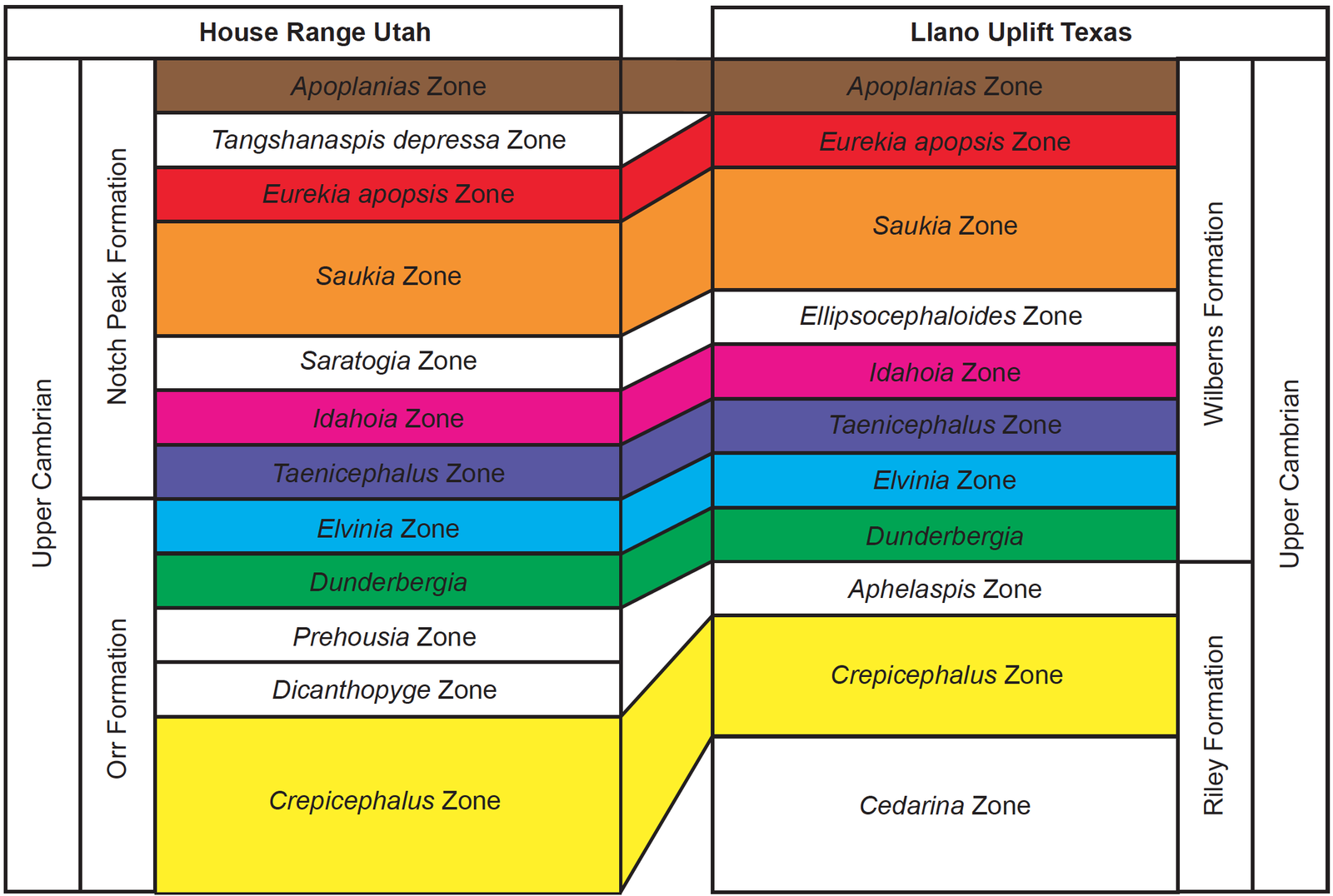
Fig. 13. Biostratigraphic data. Comparison of two suites of trilobite zones, one suite from the upper Cambrian Notch Peak Formation in Utah, and one suite from the Llano Uplift in Texas. Both suits contain eight trilobite zones that not only correlate, but correlate in their relative positions within the sediments. Adapted from Miller, Evans, and Dattilo (2012): Miller, Loch, and Taylor (2012).
Importantly, one does not have to accept uniformitarian models when interpreting these biostratigraphic data. Although the deposition of these zones during the year long Flood is unlikely, they could have been deposited in a century, or even a decade, especially if these trilobite zones represent migrating ecosystems rather than trilobite evolution. Millions of years is not required.
In conclusion, I believe these criteria supporting a real, geographically extensive and in situ pre- or post-Flood environment are quite compelling and conclusively demonstrate that an in situ growth model for the Utah microbialites is the only viable alternative. As such, I believe that these forms can be used as a proxy for other, very similar stromatolites in North America, China, and Korea. A number of solutions will now be summarized that seek to grapple with these data in other ways.
The Great Unconformity Location Solution
It will be timely here to remind the reader what all the fuss is about. Most creationists tie the onset of the Flood to the Great Unconformity, especially in North America, which separates the Cambrian from the underlying Precambrian strata. But if the Great Unconformity represents the onset of the Flood, then that means that all of the stromatolites discussed above grew during the first few months of the Flood. Clearly, this is a problem that needs to be resolved. One such solution proposes that the Great Unconformity does not underlie these stromatolites, but there are a number of problems with that solution.
The location of the Great Unconformity is best delineated by the presence of the detrital sandstones that unconformably overlie the erosive surface itself (fig. 6). The Tapeats Sandstone in the Grand Canyon is perhaps the go-to place for creationists seeking to describe, understand and showcase this underlying erosive surface. The Tapeats Sandstone and its related formations are regional in scope, extending over much of present-day North America (Miller et al. 2012; Peters and Gaines 2012; Yonkee et al. 2014). Fig. 6 clearly shows that most of the Cambrian microbialite locations described above sit stratigraphically above those sandstones and thus above the Great Unconformity. I am not familiar with the geology of Korea or China, so I cannot comment as to the relationship between those microbialites and the Great Unconformity in those locations, but since the entire Cambrian–Ordovician succession in those locations sits atop a sandstone formation that itself sits upon Precambrian basement rocks, then this correlation is entirely possible.
The correlative formations for the Tapeats Sandstone in and around my research site are known as the Tintic Quartzite in central Utah, the Geertsen Canyon Quartzite in northern Utah, and the Prospect Mountain Quartzite in southern Utah (Yonkee et al. 2014). This latter Formation underlies the Notch Peak Formation, with many exposures showing a clear contact between the Prospect Mountain Quartzite and overlying limestones (Miller, Evans, and Dattilo 2012a).
Hintze and Davis (2003, 33 and 214) have exceptionally detailed maps and stratigraphic columns that identify gneissic basement rocks well below these overlying Cambrian sediments (fig. 3). Kilometers thick assemblages of Proterozoic quartzites overlie the gneisses and the Prospect Mountain Quartzite sits on top of those quartzites. Since the 2–3 km thick succession of carbonates, including the Notch Peak Formation overlie the Prospect Mountain Quartzite, we can be quite confident that the Great Unconformity should underlie this Formation, even though the Cambrian/Precambrian boundary here appears to be conformable.
It is quite clear that many Cambrian microbialite locations in North America, and perhaps even in China and Korea, sit stratigraphically above the Great Unconformity. As such, the solution that seeks to “move” the Great Unconformity from these overlying Cambrian stromatolites is fraught with other problems.
The Allochthonous Solution
Creationist literature detailing Paleozoic/Mesozoic reefs in general often adopt an allochthonous interpretation of deposition. This predisposition most likely stems from a creationist misconception that relegates kilometers thick assemblages of stromatolites to Precambrian strata and small, unconsolidated coral bioherms to Paleozoic and Mesozoic strata (traditional Flood deposits). This distinction has obvious benefits in that small, unconsolidated coral bioherms are more easily explained as allochthonous buildups.
In recent years, however, more and more Cambrian stromatolite reefal complexes have been described in the secular scientific literature. As I have outlined above, some of these complexes have extraordinarily large geographic distributions and are composed of tightly packed individual stromatolite structures that clearly grew in place. These assemblages are much more difficult to explain allochthonously, and often defy traditional creationist reefal definitions which tend to focus on Silurian through Permian unconsolidated bioherms (Whitmore 2009). Since transport of individual reefal components is more difficult to explain in traditional creationist Flood models, some creationists may seek to interpret Cambrian stromatolites in terms of exceptionally-sized allochthonous megaclasts. I will now discuss this scenario as a possible solution.
The Cambrian rocks containing the Notch Peak microbialites represent pre-Flood environments that were pushed onto the Laurentian craton, over the Cambrian sandstones, during the Flood. In this scenario, although the individual microbialites are in situ (discussed above), the entire deposit as a whole is allochthonous. This option has the advantage of retaining a Precambrian–Cambrian Flood boundary. There are, however, severe problems with this option, most evident of which is the lack of geologic evidence supporting such a catastrophic movement of enormous land masses. Blocks that are hundreds and even thousands of square kilometers in size and perhaps several kilometers thick should leave ample evidence such as crumpling and thrust faults (Wise and Snelling 2005). Yet all the formations spanning the lower Cambrian through Upper Ordovician are conformable (Hintze and Davis 2003; Hintze, Taylor, and Miller 1988; Miller et al. 2003; Miller, Evans, and Dattilo 2012), faulting only in the Jurassic due to compressional forces and in the Tertiary due to block-faulting (Powell 1959).
In their 1998 ICC paper, Sigler and Wingerden extensively describe the megabreccias and associated sediments of the upper Proterozoic Kingston Peak Formation in the Mojave Desert region. This paper is instructive because it painstakingly details a large-scale allochthonous deposit where individual clasts reached several kilometers in diameter. If the microbialites of the Notch Peak Formation were transported in allochthonous blocks, then the associated sediments ought to compare with those of the Kingston Peak Formation. Yet the two formations could not be more different. Megaclasts associated with the Kingston Peak Formation, although retaining the “original order of parallel strata” at some locations, were redeposited as “tightly folded slabs” at others (Sigler and Wingerden 1998, 496). The Kingston Peak Formation itself contains huge breccias entrained as debris cut from the underlying Beck Spring Dolomite and the Crystal Spring Formation. Debris-flow deposits, diamictites, gravels, sandstones, turbidites, and numerous other sedimentary facies diagnostic of allochthonous transport pervade the entire Kingston Peak Formation. The Notch Peak Formation, on the other hand, lacks all of these diagnostic sedimentary facies. Yes, some wackestone, and even some packstone grain-flow deposits occur sporadically here and there, but such flows are consonant with a gradually subsiding continental margin. An allochthonous solution becomes incredulous when one realizes that the stromatolites associated with the Notch Peak Formation extend out for tens (perhaps hundreds) of kilometers in both directions, and are buried within multi-kilometers thick sequences of carbonates. Geologic evidence associated with the catastrophic deposition of such large blocks should surely be abundant.
There is also the biostratigraphic correlations discussed above (fig. 13). Multiple trilobite zones have been identified at several locations all over western and central North America. Of interest here is not the biostratigraphic “dates” associated with secular interpretations, but the relative position of the trilobites with each other. If the central craton and the large stromatolite-containing blocks are representative of Flood deposits, and the large blocks representative of allochthonous antediluvian reefal systems, then how did eight different suites of trilobites get deposited on the craton in the Flood, in the same relative order as the in situ suites in the blocks?
An allochthonous interpretation for the Notch Peak Formation must also be applied to the other stromatolite locations in North America, Korea, China, and many other places in the world, even though no evidence for catastrophic deposition exists at any of these locations. Given all of these data, this solution must be abandoned.
The Growth Rate Solution
Could Cambrian microbialites have grown during the Flood? The rate of microbialite growth has been calculated at between 5 mm a year for microbialites growing today in Shark Bay, Western Australia (Playford 1980) to as high as 36 cm a year for modern forms growing in Bermuda (Gebelein 1969) and an equally high rate of 36 cm a year for forms found in Bahamian tidal channels (Reid et al. 2000). Many factors, however, can influence this rate of growth, and so a growth rate in and of itself should not be characteristic of growth rates in general. For example, the Shark Bay microbialites seem to represent an exhausted ecosystem (Playford 1980, 73). Proximal sea level has been dropping consistently for quite some time, and many microbialites now sit within the supratidal zone, completely stranded from a prior, subaqueous existence. Since no real opportunity for further significant growth exists at Shark Bay, the very low rate of growth for these forms should not be used as a proxy for microbialites growing in more favorable environments. The modern forms growing in sand-laden channels in the Bahamas, can accrete at 1 lamination per day (365 a year at approximately 1 mm per lamination = approximately 36 cm per year), but never actually maintain this rate due to factors such as mat type, burial, lithification, and scouring by sand (Reid et al. 2000). More recently, Berelson et al. (2011) conducted an experiment on silicon microbialites growing in a pond at Yellowstone National Park. They were able to grow a microbialite from scratch and were therefore able to verify a high growth rate of 5.7 cm a year. Eagan and Liddell (1997, 302) predicted an extremely high rate of growth for ancient microbialites of between 37 cm to 60 cm a year.
These varying rates, although quite diverse, seem to reflect the environment in which the microbialites grew or are growing. Supratidal forms predictably do not really grow at all; subtidal forms that are subject to constant erosion and burial, although growing rapidly at times, tend to lose their newly acquired width and height to the erosive activity of sand. On the other hand, the forms found in Cambrian rocks by Eagan and Liddell (1997) seem to have been growing in a favorable environment—hence the high growth rates. The microbialites studied in this paper are temporally and geographically very close to those studied by Eagan and Liddell (1997), and thus serve as an environmental proxy that suggests high rates of microbialite growth.
From a creationist perspective, given extremely favorable conditions of growth, it is not implausible to consider rapid growth rates on the order of a few meters per year for upper Cambrian microbialites. Even if incredibly high growth rates such as this can be verified, however, such rates are still completely inadequate to account for hundreds of meters of accumulated microbialite growth in the Flood year. Moreover, this rapid growth is supposed to be taking place while up to 1 km of the continental shelf is being removed by Flood-generated tsunamis (see below).
I remember a fellow creationist once telling me that he believed such a feat entirely possible. Another colleague adamantly believed that all the microbialites at my research site must have grown in one year because those sediments were Flood sediments. Yet such assertions are really no different than our secular colleagues who say that dinosaur soft tissues can preserve over deep time because the bones are millions of years old. Even allowing for exceptional growth rates, it is extremely implausible that hundreds of meters of microbialites could grow in just a few months (since these sediments would have to be deposited at the beginning of the Flood). Given that many Cambrian microbialites contain sponges as well as other encrusting metazoans, a credible growth rate solution is virtually impossible.
The Abiotic Solution
Did these Cambrian microbialites form under strictly abiotic conditions? Those proposing this solution admit that biological growth rates cannot account for such microbialite thicknesses but think that abiotic growth rates might provide a solution.
Although bacterial fossils have not been found in the microbialites at my research site, many other Cambrian microbialites elsewhere were constructed by calcimicrobes, their “skeletons” now representative of the microbialite buildup itself (see above). There is also ample evidence for the biogenicity of other kinds of encrusting organisms such as the sponges outlined above (Coulson and Brand 2016). Sponges, as well as other encrusting metazoans, have also been found in Cambrian microbialites all over the world. This position becomes completely untenable when one considers Ordovician microbialite assemblages which are chiefly constructed by sponges, bryozoans, and multiple other forms of metazoan fauna.
Even if a purely abiotic mechanism were responsible, this does not alleviate the problem. The crux of the issue is not biotic versus abiotic, it is time (Purdom and Snelling 2013). In Bed 9 (see above), for example, time-dependent processes were responsible for microbialite coalescence and elongation (Coulson, Brand, and Chadwick 2016). In Bed 11 (see above), encrusting sponges, in conjunction with microbial communities, constructed 30–70 cm-high microbialites one lamination at a time. Given the overwhelming biogenicity of most microbialites, this solution should be discarded.
The Seismic Solution
Brand, Phillips, and Chadwick (2012) proposed a seismic origin for these microbialites. In their model, Brand, Phillips, and Chadwick (2012) proposed that the existence of the elongate stromatolites in the Hellnmaria Member of the Notch Peak Formation resulted from seismic vibrations during faulting (personal communication).
Subsequently, however, Coulson, Brand, and Chadwick (2016) were able to furnish a cogent growth model for the elongate forms that accords well with modern analogues (Logan 1961). In Bed 9 (fig. 2) round stromatolites at the bottom of the bed coalesce with other round forms father upsection, eventually fusing into the elongate form within the middle of the bed. These elongate forms eventually separate into round forms again a little father up-section. Coulson, Brand, and Chadwick (2016) attributed this changing morphology to the introduction and subsequent removal of a bi-directional flow regime.
There is also no modern analogue for a seismic interpretation. It is true that vibrations do produce repeating motifs given loose particles, but it seems unreasonable to assume the same sorts of processes produced the eclectic assemblage of stromatolites found in the Notch Peak Formation. Modern microbialites, although different from these ancient forms, still have much more in common and thus provide the best analogy from which to interpret the Notch Peak microbialites, and all Cambrian microbialites in general. All of these concerns are compounded given the microbialites from other locations in North America and around the world.
Once we include the ecological, environmental, and petrographic aspects imprinted within the stromatolites from the other 10 beds (figs. 2, 4, 5, 8, 9, 10, 11, and 12) and a seismic interpretation becomes untenable.
The Proposed Solution
If the microbialites did not originate within the Flood event itself, then the only other options are their growing in either the pre- or post-Flood worlds. Placing them in the post-Flood world is problematic because of the thick, stratigraphic sequences that occur directly above the Cambrian strata. These sequences range from Cambrian all the way through to Permian in age (Hintze and Davis 2003; Miller, Evans, and Dattilo 2012a). A post-Flood interpretation for the Cambrian strata would, therefore, warrant a post-Flood interpretation for these other sequences as well. Since the latter are continental in scope, a post-Flood interpretation is unlikely.
It would seem the best solution is to interpret these microbialites in terms of the pre-Flood world. This interpretation, however, has many unsatisfactory elements, the most salient of which concerns the processes that formed the Great Unconformity. If these microbialites represent in situ, pre-Flood environments, then according to the law of superposition, the Great Unconformity must have formed prior to their growth, within the pre-Flood world. This interpretation is of course not going to sit well with many creationists for at least four reasons:
- The universal erosive processes associated with the Great Unconformity fit well within a Noahic Flood model.6
- Placing the Great Unconformity within the pre-Flood world naturally entails other universal catastrophic processes outside of those associated with the biblical Flood narrative.
- The Great Unconformity and continental-wide early Cambrian marine sediments have become an apologetic Flood support within mainstream creationism. Discarding this support will seem like a step backward.
- If the onset of the Flood of Noah did not occur at the Precambrian-Cambrian boundary, then where in the geologic column is the record of that event? I propose that most7 Great Unconformity locations resulted from processes associated with Creation Week. This means that the Cambrian microbialites sitting stratigraphically above the Great Unconformity represent pre-Flood biomes. This suggestion is a tall order and has lots of potential drawbacks, but I think this position also has much to commend it and personally believe it will be most helpful in developing more robust creationist models. For the purpose of presenting a cogent position, I will first attempt to counter possible objections.
What Process Formed the Great Unconformity?
Before discussing these processes, I think it will be important to reference Austin and Wise’s (1994) paper that sought to define the on-set of the Flood using a suite of five criteria. These five criteria are:
- A mechanical-erosional discontinuity (ED) identified by regional structural analysis—probably the most significant unconformity in any given area.
- A time or age discontinuity (AD) identified by coarse sediments above the erosional unconformity containing lithified fragments of various sedimentary units found below the unconformity.
- A tectonic discontinuity (TD), found at the erosional unconformity, distinguished by substantial regional tectonic disruption, especially at pre-Flood continental margins.
- A sedimentary discontinuity (SD) consisting of a thick, fining-upward, clastic-to-chemical strata megasequence of regional to inter-regional extent defined at its base by a significant onlap unconformity.
- A paleontological discontinuity (PD) marked by an increase in abundance of fossils and the first appearance of abundant plant, animal, and/or fungal fossils (Austin and Wise 1994, 1).
In order to make a case for a later Paleozoic pre-Flood/Flood boundary, these five criteria need to be assessed very carefully. At the outset, I want to state that I mean no disrespect to these two men, both of whom have massively contributed to my own growth in creationist research. My reevaluation of these criteria are built upon my own presuppositions as to the nature of the Flood and in no way diminish the criteria if their presuppositional position is correct. I could be wrong. But until I am persuaded by science or the Bible, I believe that my presuppositional position is justified.
Regarding the first criterion, I would argue that the unconformities produced during Creation Week would either be as significant or more significant than the Flood. Many creationists have discussed the likelihood of catastrophic erosion and sedimentation associated with emergent land masses on Day 3 of Creation Week (Dickens 2017; Dickens and Snelling 2008; Snelling 2008; 2009). In my book, Creation Unfolding: A New Perspective on Ex Nihilo (Coulson 2020), I argue that the processes associated with Creation Week, although only taking six calendar days, more than likely occurred at highly accelerated supernatural rates. This would mean that the geologic work associated with the events of Day 3 might equate to millions of years’ worth of geologic work at rates experienced in the present. Snelling (2009, 469) agrees: “What is clear from the biblical record is that the perception of a human observer on the earth during Creation Week would have been that countless millions of years of earth history at uniformitarian rates had been compressed into six days of normal human experience.”
Austin and Wise (1994), however, believe that processes associated with Noah’s Flood, and not those of Creation Week, constitute the greatest catastrophe in history. As such, they believe the most significant surface of erosion in the geologic record should be equated with the events of Noah’s Flood. It is true, Creation Week regression technically occurred outside of “history” as we know it, but the sheer magnitude of Day 3 processes should surely leave its mark in the geologic record. Consider these words from Snelling (2009, 613), “In this short time span, the earth was created and established with an almost incomprehensive amount of geological work completed in order to produce as much as half or more of the geologic record.” No matter how one interprets the events of Creation Week, one thing is quite clear: The events of Day 3 not only could, but I venture to say, should have left gigantic scars in the geologic record. Given this presuppositional position, which, I trust is scientifically and biblically justified, I contend that at least one of those scars is the8 Great Unconformity.
The second criterion is again dependent on one’s presuppositional starting point. As a new creationist many years ago, I had never entertained the possibility of antediluvian epeiric or inland seas. It is not that I did not know what they were; it is just that no one in the creationist community seemed to champion such a position. As such, I just rallied along contrasting and interpreting the data as per the status quo. But as I began to see the sorts of things described above, I began to think of why such an idea was out of the question. And I could not find one, not biblically or scientifically.
I realize that Austin and Wise (1994) do not hold such a position, and as such, their criteria are going to reflect their own set of assumptions. But, if there were such things as large epeiric inland seas in the pre-Flood world, then that would affect the criteria laid out above. For example, if large continental areas were covered with huge bodies of water, then one would not expect all underlying substrate to be lithified. Some or most of it would actually be soft. This, of course, means that the onset of the Flood, although tearing down to basement rocks at some locations, may only have excavated soft micritic muds and/or clays and shales at others. This means that the second criterion would not hold for submerged continental land masses, although it would for upland areas.
The third criterion, as with the first, does not take into account processes at work during Creation Week. In fact, throughout their 1994 paper, Austin and Wise do not even mention Creation Week, which surprised me given the obvious tectonism that would have been involved. What I am saying then is that large “erosional unconformities” and/or “tectonic discontinuities” may have resulted from Creation Week processes and not those associated with the Flood. This rationale must be allowed to stand as a scientific and biblically valid possibility.
The fourth criterion is again only valid if large epeiric inland seas did not exist in the pre-Flood world. “Thick, fining-upward, clastic-to-chemical strata megasequences” would not be consistent with a transgression over already submerged continents that themselves are blanketed with thick sequences of marine carbonate muds. Now, this would not be the case for subaerially exposed continental rocks, so the criterion is a good one, but only where applicable.
The final criterion has always bothered me. The onset of the Flood is marked by: “the first appearance of abundant plant, animal, and/or fungal fossil[s]” (emphasis mine). This assumption is not limited to Austin and Wise (1994). Most mainstream creationists accept this interpretation (Snelling 2009). Even in the traditional creationist model, however, there is no reason not to expect some animal fossilization prior to the onset of the Flood. Austin and Wise (1994) assume that there was no major tectonic activity prior to the Flood, but how do they know this? These next few paragraphs are going to get a bit wordy, but I must explain myself here.
Most creationists who assume that no animal fossils should be found in pre-Flood deposits believe this because they say that the Bible does not discuss significant tectonic activity prior to the Flood. But the Bible does imply that the entire planet was subjected to a supernatural and catastrophic curse at the Fall of Adam. I think it is easy for us to forget that the judgement of God in the Flood is but one part of the greater judgement of God in the curse. God cursed every single creature and all its progeny with death. He cursed the earth and the land. According to Paul, the curse brought a universal “groaning” to the creation. Paul says:
For the creation was subjected to futility, not willingly, but because of him who subjected it, in hope that the creation itself will be set free from its bondage to corruption and obtain the freedom of the glory of the children of God. For we know that the whole creation has been groaning together in the pains of childbirth until now. (Romans 8:20–22 ESV)
Most creationists I talk to agree that this “groaning” and “bondage” includes geologic activity like earthquakes and volcanic eruptions that have been with us since the Fall, not the Flood. Taking the text at face value, I would suggest that fossil evidence for the Fall in terms of billions of dead animals should be seen throughout the geologic column, and not just in Flood rocks. Since God’s judgement against living organisms began at the Fall, I should also expect to see billions, or at least very large quantities, of dead animals in pre-Flood deposits. Yet where are they? There is not a single example of even a mollusk!
Is it not strange that in the entire Precambrian fossil record, we cannot even find a single shelled organism, let alone more complex animals that lived in the antediluvian world? Given violent earthquakes, volcanic eruptions, tsunamis, cyclones, etc., should not at least a few organisms be buried and fossilized? Given the presence of ubiquitous pre-Flood streams and rivers, couldn’t a few vertebrates be sequestered in sediments? These data, in conjunction with the existence of large sequences of Neoproterozoic sandstones and conglomerates, indicative of energetic sedimentary processes, lead me to really question the legitimacy of this traditional creationist interpretation.
The lack of even a single shelled organism in the Precambrian is better explained as a consequence of special creation in conjunction with an historical curse rather than the Noahic Flood. This solution lends more credibility to the utter disparity that exists between two suites of strata separated by just millimeters of rock.
Another important matter concerns the geologic “cone of silence” that many allude to for the purpose of precluding large scale pre-Flood tectonic activity. In other words, since the Bible does not talk about energetic pre-Flood processes, we cannot use science to fill in the gaps. Lest I am accused of reading too much science into the Scriptures, I would like to posit the following example. We find millions of fossils, for example, in Ice Age deposits. According to Whitcomb and Morris (1961, 156), there might be as many as five million mammoths buried in these deposits, not counting other kinds of mammals. Yet some overly skeptical conspiracy theorists believe there was no such event as an Ice Age, and that all of these animals died in the Flood of Noah. In the minds of these people, mainstream creationists have used science to fill in what the Scriptures do not supply. And this accusation is valid. Mainstream creationists are quite convinced that, based on the scientific evidence, these deposits consisting of millions of dead animals could not have directly resulted from a global Flood. Before this evidence became mainstream, however, these fossils were used as prima facia evidence for the Flood. Essentially, we have used science to explain a gargantuan record of fossilized mass death that is not referenced in Scripture.
Creationists that adopt the Cretaceous/Paleogene post-Flood boundary must apply the same logic to the billions of fossils found in other Cenozoic deposits. Yes, there are scientific solutions that fit with the biblical narrative, but if we are completely honest, we are using science to explain what the Scriptures do not necessarily discuss. The Bible says nothing about the death of billions of animals in post-Flood catastrophic processes. Yes, it seems to suggest very “icy” conditions in the book of Job, and depending on how you interpret Psalm 104, there may be a reference to the uplift of mountains, but generally, the Bible is completely silent about the catastrophic death and burial of billions of Cenozoic animals.
Finally, I would suggest that the Bible is actually not silent about powerful antediluvial geologic forces. This is an assumption, that, oddly enough is built on uniformitarian ideology. Most creationists agree that God’s curse upon the earth included the advent of earthquakes and volcanic eruptions, much as Paul experienced them in his day and we in ours. This is the geologic “norm.” But how do we know that the rates of geologic change experienced in the present are necessarily the same as the those experienced in the antediluvial world? This assumption is uniformitarian through and through. The geologic effects of the curse in the pre-Flood world may have looked vastly different than the effects of the curse that we see in the present. In other words, tectonic plates that moved at several meters per week, mountains that significantly changed their elevation in decades, and thick packages of sediment that filled basins in just a few years, might have been their “geological norm.” As an analogy, consider the people living just a few hundred years after the Flood. Most creationists believe that post-Flood geologic activity was quite aggressive, decreasing gradually to levels we experience today in just a few hundreds of years (Snelling 2009). Now consider some descendants of Noah’s family living at that time. Depending on where they lived, this group of people may have experienced extraordinary (to us) geologic change during their lifetimes. They may have literally seen mountains grow in just a few decades, or basins filled with sediment in just a few years. Yet for them, this was normal. This is what many Christians do today—they think that the rates of geologic change we experience in the present reflect a universal “geologic norm.” But this, of course, is uniformitarianism plain and simple.
Importantly, all Christians explain the presence of life-destroying hurricanes, volcanoes, earthquakes, and tsunamis as a consequence of the Fall, not the Flood. In other words, the Bible discusses the presence of these sometimes catastrophic forces, and their presence in the real world has a biblical foundation. Given non-uniformitarian assumptions, there is no reason to deny the same rationale for the existence of aggressive geologic processes in the antediluvian world.
In conclusion, the criteria we use to determine a pre-Flood/Flood boundary is going to depend on our presuppositional starting conditions. Secondly, the “apparent” silence of Scripture can in no way be used to preclude the presence of powerful tectonic pre-Flood processes. Creationists regularly use science to explain phenomena as long as it doesn’t contradict the Bible. This means creationists should be free to use the science as long as they can remain committed to a faithful interpretation of Scripture. This should be kept in mind with all that follows.
What about Precambrian Ecosystems?
If cratonic basement rocks were beveled almost flat during Creation Week, then that would mean that most of the sediments associated with the Precambrian were deposited in the same six-day period. Yet are there not dozens of horizons of stromatolites spanning many thousands of meters of sediment in the Precambrian? And, of course, there are.
To answer this question, I would like to turn to some Archean stromatolites. According to Purdom and Snelling (2013) and Clarey (2018), Archean rocks and stromatolites came into existence during Day 3 regression of Creation Week and so represent specially created buildups. Some of these stromatolites can be found in the Pilbara region of Western Australia within the Early Archean Strelley Pool Chert (Allwood et al. 2007). This stromatolitic assemblage, which is about 10 m thick and can be correlated for about 10 km, initially grew on a basaltic conglomerate. The stromatolites change morphology upsection from domicile laminates that encrust the basal boulders, to large 1 m thick cones and eggshell carton shapes in the middle, to small wavy laminates at the top. Small cusped forms as well as flat laminae also occur in the section. Although the biogenicity of Archean forms is often challenged, Allwood et al. (2007) makes a good case for the biogenicity of these buildups and interprets the changing morphologies in terms of a shifting intertidal environment. Snelling (2009, 633) agrees with the biogenicity of these stromatolites.
Since most creationists accept the Creation Week origin for these stromatolites, the presence of morphologically disparate microbialites in middle to upper Precambrian sequences could likewise be interpreted as Creation Week buildups. Interestingly, creationist Kurt Wise at one time suggested that upper Precambrian stromatolites were specially created on Day 3 of Creation Week due to a lack of animal fossils found in association with the stromatolites (see Wise and Snelling 2005, 23). Both Wise and Snelling now dismiss this interpretation for many upper Precambrian stromatolites based on the following logic; Precambrian stromatolites were either:
- Created as fossils,
- Created as fully functioning entities that had a purpose, or
- Developed according to normal processes in the pre-Flood world.
According to both authors, the Neoproterozoic (upper Precambrian) stromatolites of the Awatubi Member of the Kwagunt Formation in the Grand Canyon area, for example, cannot have been created as fully functioning entities during Creation Week because these buildups occur on multiple horizons. If the uppermost horizon represents the terminating Creation Week buildup, then that means the lower horizons must have been created as fossil forms which is, rightly I might add, unacceptable. According to these authors, “A fossil stromatolite does not seem to be fully functional,” an observation with which I agree.
But I think there is a fourth possibility not discussed by Wise and Snelling (2005). What if Creation Week unfolded in a time-lapse fashion? For a full discussion of this hypothesis, I direct the reader to my book (Coulson 2020). In the book, I essentially say that Creation Week microbialites serve a secondary purpose by infusing the atmosphere with oxygen. This idea is, of course, in agreement with the secular scientific community, but naturally precludes the time involved.
God’s use of secondary processes to fashion the earth during Creation Week is not a novel idea. Snelling (2009, 627) proposes a secondary origin for other atmospheric gases during Creation Week: “Regarding the formation of the earth’s atmosphere, it is also likely that, in addition to the convective circulation in the mantle bringing heat to the earth’s surface, it would also have brought gases to be released above it and accumulate as the atmosphere” (emphasis mine). Snelling proposes secondary processes for earth’s soils (which involve bacteria), the earth’s metallic reservoirs (Snelling 2008), and for the formation of its continental crusts (Snelling 2009, 628). Baumgardner (2000) proposes a similar scenario for the differentiation of the earth’s core and crust from the earth’s mantle, as does Faulkner (2016) for many other facets of the created order.
Genesis itself strongly implies that God used rapidly accelerated processes to mature the biosphere: “And God said, ‘let the earth sprout vegetation, plants yielding seed, and fruit trees bearing fruit in which is their seed, each according to its kind, on the earth’” (Genesis 1:11–12 ESV; emphasis mine). Any cursory reading of this text implies that the entire vegetated realm popped up in 24 hours! This miraculous method of achieving maturity has many examples in later sections of Scripture. Consider Numbers 17:8b, where we are told that the staff of Aaron, “had sprouted and put forth buds and produced blossoms, and it bore ripe almonds” (Numbers 17:8b ESV; emphasis mine). That God used supernaturalism to fashion a planet using accelerated “naturalistic-like” processes fits with His creative strategy—to frame a world for man. Why wait billions of years if that was His primary concern? Using these “naturalistic-like” processes to saturate the atmosphere with oxygen should, therefore, not surprise us.
In this scenario, the lower stromatolitic beds in the Awatubi Member were not created as fossils. Rather, the entire stratigraphic sequence was deposited on Day 3 of Creation Week, with each stromatolite layer serving a temporary purpose—oxygenating the atmosphere. After this purpose was served, the stromatolite layer was buried. The next horizon of stromatolites served the same purpose before it too was buried, and so on and so forth.
Importantly, these accelerated inorganic (sediment deposition) and biological (stromatolite growth) processes occurred at the same rates relative to each other. For an observer, the growth of these stromatolites would look no different than what one sees in the present, albeit at an accelerated rate (see also Snelling 2009 and Coulson 2020). This mode of quickening fulfills God’s desire to build and frame a world in literally six days, without losing any of its paleo-environmental significance. In other words, paleontologists can understand these sediments and stromatolites in terms of processes which in turn leads to a real environmental interpretation. Christian paleontologists, however, must not confuse bulk process with time. That is because God used process to create a mature world. Christians must believe what God has said in His Word and refrain from applying uniformitarianism to historical counterparts when thus informed. This, of course, is an act of faith that accords with God’s revelatory design (Hebrews 11:3).
Our secular colleagues rightly scoff at such absurdity. How would the master of the feast in John chapter 2 have reacted had he been told that his Yarden Katzrin Red was pulled from thin air! In his mind, and in the mind of the other guests, Jesus’ wine grew on vines, was crushed in a wine press, and fermented in a vat, perhaps for years. You see, the wine was natural. The miracle was in the making of the wine. And that requires faith. Yes, the earth is a lot bigger than 180 gallons of wine, but the principle is the same. And yes, the wine was created without any process whatsoever, but there are other examples in Scripture where “naturalistic-like” processes were at work. There is the withering of the fig tree in Mark 11, or the restoring of atrophied retinal cells in John chapter 9. Of course, the rapidly maturing vegetated realm from Genesis 1 is a perfect example of these “naturalistic-like” processes. But whatever the miracle, whether wine, fig tree, fish, plants, or baked bread, the end result is the same—natural objects that retain natural histories. Really, if God can bring a man back from the dead, then can He not construct a universe in six calendar days? Of course, accepting this requires faith, and that of course brings us right back to Hebrews 11:3 ESV: “By faith we understand that the universe was created by the word of God.” I think this verse is better appreciated when stated negatively: “Without faith, we cannot understand that the universe was created by the word of God.” A completely secular view of our planet’s origin requires no faith and is, therefore, erroneous—at least according to Scripture. In the model presented here, the Precambrian should be viewed like Jesus’ wine—natural in every way, but specially and supernaturally framed. Yes, a seeming oxymoron, but one that accords fully with Scripture requiring faith to accept.
In conclusion, whether or not one accepts these accelerated, “naturalistic-like” processes doesn’t change the reality of a morphologically diverse Archean microbialite assemblage that was specially created and subsequently buried during Creation Week. Its existence sets a precedence that allows for other similar examples in the rest of the Precambrian.
What About the Ediacaran Biota?
The creation and subsequent burial of specially created Precambrian ecosystems, such as microbialites, naturally implies the death of the microbes involved. Most creationists see no problem with such a scenario, and will even include protists, fungi, algae, plants, as well as other kinds of basic eukaryotic organisms (Wise and Snelling 2005). Animals do not appear in Paleo-Mesoproterozoic rocks, and so creationists should not object to the special creation of these rocks. But what about the enigmatic Ediacaran Biota of the Neoproterozoic Ediacaran Period? (This would also be true for putative sponges that have been found in rocks of similar age9). Do these organisms represent “living” entities in the biblical sense? Most scientists give that distinction to organisms that move, metabolize, breath oxygen, grow from a blastula, and reproduce sexually. We label such organisms “animals.” Are Ediacaran organisms animals? Since some of them seem to have been motile (Chen et al. 2019), the answer to this question is most likely in the affirmative, but some doubt persists:
Although many of these organisms may represent animals (Metazoa), few share any synapomorphies with extant metazoan clades and thus may represent extinct groups with no modern representatives . . . Consequently, the position of the Ediacara biota in the context of the late Neoproterozoic-Cambrian rise of animals remains poorly understood. (Darroch et al. 2018; emphasis mine)
More than likely, then, these organisms were biblically “alive,” but their fossils would need to be found in strata associated with the Creation Week before a conflict arises. As it turns out, most of these fossils are found below the Great Unconformity (Corsetti and Kaufman 2003), but this should not automatically necessitate a Creation Week origin (if in fact, the Precambrian is representative of Creation Week).
Traditionally, the onset of the Flood has been tightly associated with the Great Unconformity (Whitcomb and Morris 1961). Snelling (2009, 710), however, has proposed a boundary below the Ediacaran Period (635 Ma), at surfaces of erosion below that of the Great Unconformity for the onset of the Flood:
However, in recent decades, unusual multi-cellular animal fossils, the so-called Ediacara fauna, have been found in late Neoproterozoic sediments below the Precambrian/Cambrian boundary in those regions where the relevant portion of the geologic record has been preserved and exposed to view. Furthermore, stratigraphically below those fossils, thick conglomerate units have been found that have been called diamictites, and interpreted as glacial deposits known as tillites, but which can equally be regarded simply as breccias that are consistent with a major tectonic disturbance. These units correspond to the Sixtymile Formation in the Grand Canyon, and the Kingston Peak Formation in the Mojave Desert region of southern California. They are also found in the Wasatch Mountains of central Utah, the MacKenzie Mountains of western Canada, in the Adelaidean and adjoining basins of southern Kalahari craton of southern Africa . . . Therefore, these breccia units could well mark the tectonic upheaval that is to be expected for the onset of catastrophic plate tectonics . . . [at] the commencement of the Flood. The Precambrian-Cambrian boundary above the Kingston Peak Formation is especially interesting. The contact between the Stirling Quartzite and the Wood Canyon Formation not only grades smoothly from one facies into the next, but it lacks evidence for large-scale tectonic and sedimentary upheavals (Hagadorn and Waggoner 2000; Snelling 2009) (fig. 14). As it turns out, even the Ediacaran fossil assemblages associated with the Stirling Quartzite blend into the lower member of the Wood Canyon Formation (Hagadorn and Waggoner 2000) (fig. 14). On the basis of these fossils, the secular scientific community has placed the boundary between the Precambrian and the Cambrian somewhere in the lower member of the Wood Canyon Formation.
This boundary is not very appealing from a traditional Flood perspective because it does not fit the criteria laid out in Austin and Wise (1994) (see also above). As such, Snelling has placed the onset of the Flood in the Mojave Desert region at the base of the Neoproterozoic Kingston Peak formation. Snelling’s rationale is associated with the thick breccias, themselves evidence of large-scale tectonic activity that lie at the base of the Kingston Peak Formation (fig. 14). The latter is separated from the Cambrian Wood Canyon Formation (and thus the Great Unconformity) by the Noonday Dolomite, the Johnnie Formation, and the Stirling Quartzite (Corsetti and Kaufman 2003) (fig. 14). Consequently, all of these sediments and the associated Ediacaran fossils are placed squarely within what creationists usually identify as “Cambrian” sediments. This means that these Ediacaran organisms lived in post-Creation Week environments.
Moving Precambrian boundaries around like this should not be considered too unorthodox. Apart from the Ediacaran fossils, there are no solid biostratigraphic indicators that help constrain “ages,” leaving just a few geochronological methods from which to accomplish this task. Karlstrom et al. (2020), for example, moved the Sixtymile Formation in Grand Canyon area to the base of the Tonto Group on the basis of detrital zircon U-Pb dates. It is now considered a Cambrian formation.
Fig. 14. Mojave Desert stratigraphic section spanning the Precambrian/Cambrian boundary, adapted from Corsetti and Kaufman (2003). The vertical dashed line corresponds to the maturing of various Day 3 sedimentary environments (see Coulson 2020 for a discussion on “dynamic creationism”). The line is dashed to indicate an imperceptible evolution of Creation Week environments with those of the post-Fall world (the virtual lack of geologic activity in the pre-Fall world prevented the accumulation of large sedimentary sequences. Creation Week and the post-Fall world are similar in that both experienced active tectonic regimes). The production of Creation Week rocks could potentially continue all the way to the base of the dotted line (the pre-Fall world left no geologic record). The dotted line corresponds to rocks that contain the first animal fossils and is thus indicative of animal death, marking a definite boundary with the post-Fall world. The Great Unconformity passes through relatively “normal” rocks in the lower Member of the Wood Canyon Formation. This Great Unconformity location is insignificant when compared to the Great Unconformity location in Grand Canyon. In my model, the latter Great Unconformity was likely fashioned during Creation Week while the former was made at some early stage in the pre-Flood world (Snelling 2009). This means that the Ediacaran Fauna at this location lived in what creationists identify as “Cambrian” time. These Ediacaran and other Cambrian organisms, such as trilobites, were either moved or migrated from their “partitioned” pre-Fall environments to this location and fossilized. The solid line represents tectonically sluggish environments (large inland seas) where thick sequences of carbonates accumulated in the antediluvian world. Tectonic activity increases over time culminating in the Flood-proper as outlined in Genesis 6–9 at some point farther up in the geologic column.
So, what does all of this mean? It may mean that the Ediacaran Biota represented “Cambrian” organisms that were geographically partitioned from other geographically partitioned Cambrian organisms (like trilobites), and were the first to be buried because of some unique, and as yet unknown, environmental conditions associated with that partitioning. This hypothesis has some validity given the grading of Ediacaran fossils into the lower member of the Wood Canyon Formation and given documented cases of Ediacaran fossils that have stratigraphic ranges even up into Cambrian strata (Hagadorn and Waggoner 2000, 355).
What About Thick, Geographically Widespread Lower Paleozoic Sedimentary Sequences?
Some Lower Paleozoic sedimentary sequences have thicknesses of more than 4000 m (Hintze and Davis 2003) and are geographically widespread. Cambrian carbonates in North America, for example, are continent-wide and often even more extensive when continental shelves are factored into the equation. Traditionally, thick sequences of Paleozoic strata have been interpreted as Flood rocks by creationists (Brand 2007; Snelling 2009). But if Lower, and especially Middle Paleozoic rocks represent real pre-Flood ecosystems, then how do we account for swaths of largely marine rocks that have been deposited upon apparently “dry land” without the aid of the Flood?
I propose that continents containing carbonate-bearing sediments and thick sequences of microbialites (for example, North America) be interpreted much as they are in the secular scientific literature—as large, shallow-water inland seas. Cambrian ecosystems then, are not, for the most part, allochthonous, but represent pre-Flood environments where Cambrian-specific fauna flourished. Ordovician ecologies most likely first flourished in off-craton oceans and were pushed onto the North American craton during post-Fall but pre-Flood tectonic activity.
This, of course, would require a plate tectonic regime that differed from that in the present, where rates of plate movement were more rapid, perhaps working at meters of displacement per week, but not as rapid as those working during the Flood where velocities apparently reached meters per hour (Baumgardner 2018). Snelling (2009) acknowledges that geologic processes operating today were different than those operating during the Flood which were different again from those operating in the pre-Flood world:
Thus, we could expect that the features and strata of the pre-Flood rock record are different from the rock record produced by the Flood, with features even different from what is being produced by geological processes operating today. It is even unclear whether uniformitarian rates of geological processes, or even exactly the same geological processes, were operating during the pre-Flood period as those operating today. (Snelling 2009, 470; emphasis mine)
Consider the North American Taconic orogeny as an example. According to Stanley and Luczaj (2015), the North American craton was dominated by a vast inland sea in Cambrian time (fig. 15). During Middle Ordovician time, small volcanic islands known as island arcs began to accrete onto the then south-eastern side of the craton. As these arcs pushed against the continent, mountains began to form (the beginnings of what we today call the Appalachians). Due to forces pushing the ground up, other forces began to push the ground down on the western side of the “Appalachians.” This depression is known as a foreland basin. As the mountains rose up, sand, shale, and mud (flysch) from the growing mountain were deposited in the foreland basin as turbidites. All these sediments eventually filled the basin leaving a thick clastic wedge over much of present day eastern North America (fig. 15).
Fig. 15. Proposed North American environment during antediluvian time. In this low-resolution approximation, most of North America was an inland sea. Upland areas existed at other locations. Adapted from Stanley and Luczaj (2015).
Creationists must disregard the time over which these events are believed to have occurred, but there is no need to disregard the sequence. Given a non-uniformitarian plate tectonic regime, all these sediments could have been deposited very rapidly, in centuries, decades, years, or even months.
In a creationist model, microbialites grew in a North American shallow-water inland sea (fig. 15). This environment, unequalled in the modern world, would have provided perfect conditions for calcium carbonate (lime mud) to precipitate directly from the water column (Robbins, Tao, and Evans 1997). Given the warm climate, shallow water, water chemistry, and large geographical distribution, it would not take long for currents to move large quantities of micritic lime mud from the central craton to shelf margins where it would accumulate as kilometers-thick sequences. During times of quiescence, microbialites would grow, during times of subsidence or rapid deposition, microbialites would be buried.
During Ordovician time, and due to rapid plate movements and island arc collisions, growing upland areas on the Eastern Seaboard deposited clastic sedimentary rocks over the once thriving carbonate platform, with some of these deposits reaching the far mid-west (fig. 15). Interestingly, the events associated with the Taconic orogeny closely coincide with the Great Ordovician Biodiversification Event or GOBE as it is known. This event is hailed by the secular scientific community as the most dramatic evolutionary radiation of marine life in history (Stanley and Luczaj 2015). From a creationist perspective, GOBE might better be explained as a biological relocation. Accretion of island arcs onto the east coast would also have caused the mingling of cratonic and adjacent ocean-basin marine life. It would not take long for these new forms of marine animals to make their way across the North American craton, diversifying, and replacing some of the Cambrian ecosystems.
But What About Sequences That Were Catastrophically Deposited?
Such a model is also consistent with other Early Paleozoic Geology. The St. Peter Sandstone, for example, is a vast sheet of nearly pure Ordovician sand that blankets the North American craton, extending laterally from Minnesota to Arkansas and from Illinois to Nebraska and South Dakota in the west. The deposition of this pure sheet of quartz sand over such a large area does not accord well with secular interpretations due to the absence of channels and the purity of quartz sand. There is no modern analogue that can explain how 650,000 km2 of pure quartz sand can be deposited over a continent in a sheet-like fashion.
Huge bodies of sand such as this could easily have been transported across the North American epeiric ocean given the tectonic regime envisioned in the model proposed here. It is true, this sandstone can also be interpreted in terms of the Flood-proper. Either model works and both fit within a biblical paradigm that testifies to earth’s catastrophic past. The model outlined here, however, allows for catastrophic deposition, such as the St. Peter Sandstone, but also allows for extended environmental accretion of sediment when geologically required—Cambrian stromatolites.
“In-between” depositional environments are also best explained using this model. The approximately 200,000 km2 Upper Ordovician Kope Formation covers large areas of Ohio, Indiana, and Kentucky. The formation has an average thickness of about 80 m and is composed of about 80% silts and muds intercalated with hundreds of thin (decimeter-sized) skeletal limestone beds. The Kope has been interpreted as part of a huge siliciclastic-carbonate ramp, and the limestone beds are interpreted as tempestites (storm deposits).
This formation has special interest for me as it became the topic of my senior capstone project while in college. As I studied the literature and spent time in the field, it quickly became apparent that these deposits did not really fit the secular interpretation. The chief problem, and the one I discussed in my senior project, was the exceptional preservation of skeletal fossils, especially the shells in the laterally extensive shell beds. The shells lacked evidence for intense bioerosion and dissolution. Almost all of the shells I looked at only had one or just a few holes bored into them, with shell ribs and other diagnostic features in relatively good condition. The taphonomic literature, however, suggests that unless shells are permanently sequestered in the fossil record, they will become fodder for bioeroders such as snails and various other epibionts, eventually breaking down in just a few decades. Yet huge storms supposedly excavated the seafloor for 5 million years during the deposition of the Kope. How could these shells be incessantly recycled for millions and millions of years and yet remain in good, and sometimes exceptional condition? Yes, breakages are common and sometimes the skeletal debris is preserved as hash, but the degree of bioerosion is still relatively low. This kind of preservation really only makes sense given a maximum of just a few hundred or a few thousand years of recycling. It also turns out that obtrusion (smothering) deposits are common in the mud layers and have been interpreted in terms of rapid deposition (Hughes and Cooper 1999). Some of these obtrusion layers can be traced for tens of kilometers!
So, catastrophic Flood? Well, not so quickly. I also observed that the Kope possessed dozens and dozens of hardgrounds that were infested with Trypanites borings. Some of these hard grounds had bryozoan colonies growing over them. Yet bryozoans are an exceptionally slow-growing invertebrate. These data precipitated two conflicting interpretations. It seemed quite obvious that the Kope Formation could not represent 5 million years of autochthonous deposition, but it seemed equally true that it could not be interpreted as a fleeting allochthonous Flood deposit. I still remember thinking that all the data could be interpreted parautochthonously (in-between autochthones and allochthonous) in as little as just a few decades, and in fact that seemed to warrant the most parsimonious solution. But nothing in my paradigm allowed for such “long” depositional environments.
The model proposed here allows for various depositional interpretations that all fit within a biblical paradigm. Some deposits are autochthonous, requiring many decades or centuries of deposition (Cambrian stromatolites), some are allochthonous requiring days, weeks, and or months of deposition (St. Peter Formation), while still others are parautochthonous requiring a few years or decades at most (Kope Formation).
Isn’t pre-Flood Catastrophism at Odds with Historical Young-earth Creationism?
A vibrant, pre-Flood, plate tectonic regime is, of course, at odds with most modern creationists, but not so with those of the nineteenth century. Scriptural Geologists, as they were known, were a diverse group of scholars, scientists and/or ministers who believed in the literal interpretation of Genesis. They were much like modern young-earth creationists (YEC), except they tended to accept the science of the day without too much reservation. Speaking of Scriptural Geologist Andrew Ure (1778–1857), YEC and historian Terry Mortenson says:
When Adam and Eve sinned, God cursed the earth, one effect of which, in Ure’s theory, was a long series of localized convulsive events all over the more thinly crusted ocean bottom, which culminated finally in God’s judgment of a global Flood. During this antediluvian period of 1,600 years, the regular pattern of fossiliferous secondary and tertiary strata was formed on the ocean bottom, as basaltic eruptions agitated the seas causing partial destructions of the land and its inhabitants and local elevations of parts of the seabed. (Mortenson 1998; emphasis mine).
Ure did not know anything about plate tectonics, but he understood geology, and he understood that great forces were required to push trillions of tons of sediment onto the continents. Imagine how excited Ure would have been to know that he successfully predicted the existence of such a force! Importantly, Ure believed that fossiliferous rocks were deposited before the Noahic Flood. This was not an uncommon position for early to mid-nineteenth century Scriptural Geologists, many of whom believed that the Lower Paleozoic rocks were deposited in the antediluvian period (Johns 2016). By the 1860s, however, and most likely due to the advent of Charles Darwin’s On the Origin of Species in 1859, young-earth creationism and its corollary, the global Flood, fell out of favor in academic circles.
These ideas were revived in the early twentieth century by YEC George McCready Price (Brand 2007), and later by John Whitcomb and Henry Morris in their successful book: The Genesis Flood (1961). Importantly, Price, Morris, and Whitcomb all believed that most of the geologic column was deposited by the Genesis Flood and used the term “Flood Geology” to describe all fossil-bearing sequences (Brand 2007; Whitcomb and Morris 1961). This perspective remains solidly entrenched within mainstream young earth creationism.
Although we owe a great debt of gratitude to these early founders of modern creationism, we must, however, as with all scientific theories, assess the usefulness of the term Flood Geology. Embracing Flood Geology, sensu stricto, does not allow for the deposition of fossil-bearing sedimentary rocks outside of the Flood year. This means that no animals were fossilized during the antediluvian and/or post-Flood periods. As Brand (2007) astutely observes: “If we make the extrabiblical assumption that all fossil-bearing geological deposits began at the flood, then there are many situations where we have no choice but to interpret entire rock formations as deposited in hours, days, or a few months at most.” In other words, creationists lock themselves into an “extrabiblical” system that will not allow for other fossil-bearing sedimentary mechanisms.
As such, Brand (2007) suggests replacing the term “Flood Geology” with “Wholistic Geology.” Quoting from Brand: “‘Wholistic Geology’, in contrast with ‘Flood Geology,’ . . . attempts to explain the geologic column by taking into account potential geological activity during the whole history of the earth rather than restricting it to only the Biblical year of the flood.” A major tenet of this less restrictive term allows for the deposition of fossil-bearing sequences before the Flood, in the antediluvian world. Brand says: “There may have been extensive geological activity before and after the Flood. The geologic column contains fossils and sediments produced by the Flood, and also those produced by processes acting before and after the Flood.”
An objection might be raised based on Genesis 7:11 (ESV), “In the six hundredth year of Noah’s life, in the second month, on the seventeenth day of the month, on that day all the fountains of the great deep burst forth.” Yet this verse clearly says that it is the “fountains” of the great deep that burst forth. Snelling (2009) believes that these “fountains” may have resulted from superheated jets of water that themselves resulted from rapidly moving diverging plate boundaries. This could only occur, however, given exceedingly rapid plate movements (meters per hour) and would be unlikely given slower plate movements of a few meters per week in the pre-Flood world. The text implies that all the fountains of the great deep broke forth on a single day. In my model, plates moved at faster rates than today, but much slower than at the onset of the Flood. In God’s providence, the plate tectonic regime set in motion at an earlier antediluvian stage, became more intense over time, culminating in the climax recorded in Genesis 7:11. Brand (2007) acknowledges this possibility, and even Snelling (2009, 694) says:
Because this cold rock [Creation Week mantle rock] would have been gravitationally unstable, held in check only by the resistance of the phase boundary, the earth could be viewed as being on the knife edge of catastrophe. Very little would have been required to lose it on a trajectory leading to the Flood cataclysm. Perhaps that release occurred at the time of the Fall and was so subtle that it was undetectable at the earth’s surface (emphasis mine).
Snelling (2009) clarifies that tectonic movement at the earth’s surface was “undetectable” during antediluvian time, but he may have said this to explain the virtual absence of metazoan fossils in what he interprets as pre-Flood rocks. Many creationists believe that similar geologic upheavals, including rapid plate movements, occurred in a descending crescendo for perhaps hundreds of years subsequent to the Flood (Snelling 2009, 613).
When was this post-Fall, pre-Flood plate tectonic regime implemented? I think the two most likely possibilities are ether at the Fall and as a result of God’s curse on the earth, or 120 years before the Flood when God first informed Noah of his plans, “Then the Lord said, ‘My Spirit shall not abide in [a] man forever, for he is flesh: his days shall be 120 years [in other words, after 120 years, man will be destroyed]’” (Genesis 6:3 ESV).
Finally, what geophysical mechanisms drove this pre-Flood tectonic regime? I am not exactly sure. Incorporating two different geophysical forces to account for two different models of plate movement (Flood/post-Flood) is problematic enough (Horstemeyer and Baumgardner 2003); adding a third (pre-Flood) only compounds the problem. Having said that, it is significant that huge motile tectonic plates exist in the real world, as does their potential for moving at substantial velocities (Baumgardner 1990; Horstemeyer and Baumgardner 2003). This is promising and means that such modeling is scientifically feasible if one rejects a uniformitarian-only philosophy. It is also possible that these plate movements were related to accelerated radioisotope decay—the heat produced acting as a catalyst for mantle and crustal plasticity.
The earth possesses a natural, catastrophic strata-depositing system. This is a fact. How that system operated at differing non-uniformitarian rates, however, requires rigorous modelling. Such profound changes to natural systems, although difficult to imagine, is entirely consistent with the Bible, and even science. A lesson from the life of Stephen Jay Gould is instructive:
On a fieldtrip in my freshman geology course, my professor took us to a travertine mound and argued that the deposit must be about 11,000 years old because he had measured the current rate of accumulation and then extrapolated back to a beginning. When I asked how he could assume such constancy of rate, he replied that the fundamental rule of geological inference, something called the “the principle of uniformitarianism” permitted such inferences because we must regard the laws of nature as constant if we wish to reach any scientific conclusions about the past. This argument struck me as logically incorrect, and I pledged myself to making a rigorous analysis of the reasons. (Gould 2002, 44; emphasis mine)
The world’s current uniformitarian perspective is entrenched in our present experience. We just do not see much happening geologically speaking. But imagine a history that is non-uniformitarian. Christians should fully accept God’s supernatural intervention in history, but due to its misuse, ought only to apply such intervention when expressly told in Scripture. Christians should expect, therefore, God to have supernaturally intervened when He cursed the world. I propose that some aspects of God’s curse are related to the onset of plate tectonics. This would make sense since many natural disasters, including earthquakes, volcanic eruptions, tsunamis, and flooding (forces that must have been at work in the pre-Flood world), are directly correlated with moving, subducting, and diverging crustal plates. I contend that although God used moving plates to construct the original crust during Creation Week, this movement ceased in preparation for the creation of the biosphere, including man, only to be reactivated soon afterwards as a result of God’s curse. It is possible that the supernatural heating of the earth’s plates at the Fall, and the reactivation of global tectonics was the catalyst God used to bring about the global Flood of Noah 1,600 years later. This allows for a single, large-scale supernatural act that had non-uniformitarian flow-on effects (in light of God’s sovereign knowledge). In this model, and as stated above, tectonic activity became more intense over time, culminating in the climax recorded in Genesis 7:11). Geologic activity did not cease after the Flood, but instead gradually decreased in intensity over hundreds of years.
In terms of geologic work, the overall scenario might look a bit like a bell curve10 with the Flood occurring somewhere at the top of the curve. Notice that this model allows for an exponential increase and an exponential decrease in energy. Christians should have no problem with this kind of non-uniformitarian model. Peter reminds us of this very thing in 2 Peter 3:3-6 (ESV):
Knowing this first of all, that scoffers will come in the last days with scoffing, following their own sinful desires. They will say, “Where is the promise of his coming? For ever since the fathers fell asleep, all things are continuing as they were from the beginning of creation.” For they deliberately overlook this fact, that the heavens existed long ago, and the earth was formed out of water and through water by the word of God, and that by means of these the world that then existed was deluged with water and perished.
Importantly, Peter is not asking his opponents to don boots, hat, and pick, travel to the nearest quarry and study the strata! He is rebuking them because they do not believe the account as it is described in the Scriptures! In other words, they do not have faith. They are completely entrenched in their uniformitarian experience of the present and think that God’s judgment will, therefore, never come. Peter is essentially saying that believers should reject a uniformitarian view of the world that is solidly entrenched in our current experience, and should instead accept the biblical accounts that discuss God’s judgement in terms of geologically non-uniformitarian processes. This mindset, of course, can only be believed. Now we know what Peter meant when he called such unbelievers “scoffers.”
Sketching a Geological Model
I will now provide a geological sketch of earth history from a creationist perspective. For the purpose of simplicity, I will only consider North American geography, and I will only include events starting at Creation Week and ending in the Late Ordovician.
During Creation Week, God used supernatural rates of change to mature and frame the earth. The crust of the earth was shaped by its emersion from the primeval oceans. Likewise, the vegetated realm grew from soil and matured in a single day. Since all rates everywhere were accelerated, yet kept constant with each other, the whole effect would look something like a time-lapse movie where real environments would “evolve.” Oxygen-producing stromatolites grew on primeval micro-continents only to be buried by sand, shale, and mud, as those micro-continents coalesced to form larger cratons. Extensive surfaces of erosion were cut into crystalline basement rocks, as well as across thick sequences of newly deposited sedimentary rocks flanking immature continental shelves. Some of these larger continental platforms were inundated by shallow oceans where microbes successfully generated carbonate sediments and built stromatolites on stable continental margins (the carbonate factory). Since Creation Week was tectonically dynamic, these real marine environments were occasionally buried by mobilized clastic muds, sandstones, conglomerates, and breccias. A vertical cross-section through a suite of Creation Week rocks would reveal an eclectic sequence of unconformities, sedimentary rocks, and stromatolites (fig. 15). More than likely, this sequence would not contain any “anomalies” indicative of supernaturalism (Coulson 2020). Creation Week “plate tectonics” ceased on Day 3 of Creation Week.
Those continents (North America) covered with water were filled with specially created organisms on Day 5. Unlike the assemblages we see in Cambrian and Ordovician rocks today, these pre-fall organisms were separated and restricted to specific locations, much like in a modern zoo. By the end of Day 6 the earth was fully matured, and terrestrial animals roamed upland areas. Most Precambrian strata was completely deposited and lacked metazoan fossils because nothing had yet died.
Because of man’s sin, God cursed the ground (Genesis 3:14). This included a reanimation of a new kind of plate tectonic regime, different from that operating during Day 3 of Creation Week. Earthquakes, volcanoes, and multiple other plate tectonic related disasters began to affect all life on earth. The curse also affected the biological realm and carnivory was introduced into the biosphere. Ediacaran organisms most likely represented pre-fall assemblages that, for whatever reason, could not tolerate post-fall conditions (perhaps they could not tolerate large volumes of mobilized clastics?). Their fossils are thus restricted to the first fossil-bearing sediments (fig. 15). Thorns, thistles, flesh-ripping teeth, and other defensive-offensive specializations were also introduced, perhaps facilitated by built-in genetic instructions (Genesis 3:18 seems to suggest that something like this occurred in the plant kingdom).
The Cambrian fauna, now programmed for vanity, begin to kill and be killed. Due to the onset of plate tectonics, island arcs were accreted onto the east coast of North America. Clastic sediments damped out the Eastern Seaboard’s carbonate factory, and eventually, these sediments reached the far mid-west (fig. 15). Ocean life, partitioned as it was prior to the Fall, was now liberated from its original environment and transported onto the craton. Life on the North American craton was greatly affected by the changing conditions and many species became extinct, while others thrived.
Where to Place the Pre-Flood/Flood Boundary
Given the model outlined above, the onset of the Flood would have been less destructive when inundating already submerged continents. On the other hand, exposed land masses would have felt the full brunt of large bodies of water moving across their surfaces. These combined processes would have crippled the global carbonate factory and eroded large quantities of siliciclastics. It is proposed, therefore, that creationists should be looking for a divide in the rock record that separates marine deposits, such as limestones, from regional scale terrigenous sedimentary sandstones and conglomerates (not including Creation Week siliciclastics, some of which were remobilized during Cambrian time). Such a divide seems to occur starting in Carboniferous–Permian rocks where carbonate production (using burial flux as a proxy) drastically falls to near zero (Peters and Gaines 2012). Carbonate production picks up again in the Late Jurassic, but never again reaches pre-Permian levels.
This might mean that other Paleozoic organisms, such as those found in Silurian or Devonian rocks, represent other pre-Fall partitioned environments that were originally placed farther out in pre-Fall oceans. The same plate tectonic regime that mingled the Ordovician ecosystem with that of the Cambrian, could, at some later antediluvian stage have deposited the Silurian and then the Devonian systems over the Ordovician ecosystem.
Moving the Flood Boundary: Let’s not Throw the Baby out With the Bath Water
At this point, I want to stymie possible objections for moving the pre-Flood/Flood boundary. Some may object by appealing to the “baby and the bathwater” analogy: “If we keep moving the Flood boundary then before long, we won’t have one!” This knee-jerk reaction has some historical basis. In the nineteenth century, many old-earth creationists (OEC) kept moving the boundary farther up in the geologic column (Johns 2016). By about the mid-nineteenth century, the entirety of the Flood was placed into the Quaternary sand and gravel deposits known collectively as the “Diluvium.” Eventually, the evidence for the Flood disappeared completely into what nineteenth century minister John Fleming, and later Charles Lyell called, the tranquil flood theory (Johnson 2011). The tranquil flood theory concedes to the universal character of Noah’s Flood, but denies that any geologic evidence exists attesting to the Flood in the rock record. Hence the descriptor, tranquil.
Almost all11 Scriptural Geologists (contra the OEC), however, believed that geologic evidence supported the catastrophic nature of the Flood: “Unanimously scriptural geologists considered the Deluge to have been universal (that is, global), having left a catastrophic impact upon the earth’s strata and thus was not a tranquil effect” (Johns 2016). Those who accuse others of taking creationism back to the tranquil flood theory usually forget that it was the OEC that capitulated, not the YEC! As with the Scriptural Geologists, I believe that a biblical perspective must incorporate a catastrophic global Flood that left significant evidence in the geological record.
I would also caution creationists from dogmatically setting biblical limits on the scientific evidence. That the Flood was global in scope is a theological fact. Applying that fact to specific layers within the geological record, however, must almost always be treated as a scientific endeavor, not a theological one. I say almost because the Bible strongly implies that animal death was a consequence of the Fall (Genesis 1:29–30). Creationists should, at the very least, assign animal-bearing fossiliferous rocks to post-Fall time. This, however, ought to be the only exception. Deciding which rocks formed during the Creation Week, the antediluvian period, the Flood, and/or in the post-Flood world must, however, be considered in a mostly scientific context.
This also means that creationists should not accuse other creationists of erroneous theology based on their modelling of earth’s geologic past. As an example, a creationist could potentially ascribe a single geological Epoch to the Flood as it is described in Genesis 6–9 and still remain faithful to the biblical evidence. Some may object by citing Genesis 7:20–21 (ESV): “The waters prevailed above the mountains, covering them fifteen cubits deep. And all flesh died that moved on the earth, birds, livestock, beasts, all swarming creatures that swarm on the earth, and all mankind.” Shouldn’t rocks associated with the Flood contain most of the earth’s fossil-bearing rocks? I believe the answer is categorically, no.
First, this judgment extends only to land-dwelling organisms. God does not say anything about the death of marine animals. Most marine organisms may be very much alive or very much dead and buried before we even get to this point in the biblical narrative. The text does not even preclude the prior death of land-based animals. We must not forget that the Flood is but one smaller part of God’s greater judgement—the curse. Prior to the Flood, land-based animals had been dying for 1,600 years! Some of these might even have died in catastrophic tectonic events. We simply do not know because the Bible is silent on the issue. All we know for sure is that the remaining land-based animals and man were judged with a second judgement—the Flood.
Second, some creationists (Robinson 1996) have proposed that most of the rocks and fossils belonging to the Mesozoic and Cenozoic eras may constitute post-Flood assemblages. Accordingly, huge fossiliferous deposits of Mesozoic and Cenozoic vertebrates represent mainly extinct post-Flood organisms. Although I disagree with this hypothesis, there is nothing unbiblical as to its claims.
Third, this kind of restriction for land-based animals that lived prior to the Flood must also be applied to the mass deaths of land-based animals that lived after the Flood. Yet as discussed above, the Ice Age was an almost global catastrophe that killed many millions of animals. Many other animals died in other Cenozoic catastrophes. One cannot use the Bible to support a position without being consistent.
Finally, some may cite other reefs in other geologic periods. If we move the boundary based on Cambrian reefs, then what do we do when we get to reefs even higher in the geologic record? This is a great question and one that is difficult to answer. Yes, other reefs exist, and each suite of reefs needs to be addressed and interpreted on its own merits.
Thick assemblages of microbialite-bearing reefs only occur in the Precambrian-Ordovician. Having said that, the Silurian and Devonian strata do contain some rather large microbialite-bearing coral reef assemblages such as that found in the Canning Basin of Western Australia (Wood 1999). The Middle Devonian through end Permian, however, is mostly known for largely unconsolidated “mud-mounds.” Microbial communities resurge again in Lower Triassic rocks. For whatever reason,12 these rocks tend to have significantly more microbial buildups than at any other geologic period since the Cambrian. Even so, most of these buildups are typically only a few meters thick and only occur at a few stratigraphic intervals (Woods 2009). It is conceivable that these buildups could have grown during the Flood.
More problematic are the coral reefs that occur in the Upper Triassic rocks of Italy and Austria, and the Upper Jurassic rocks of southern England (Wood 1999). These reefs will require careful creationist research. It is entirely possible that these Upper Triassic and Upper Jurassic coral reefs represent autochthonous Mesozoic marine environments that were separated from Paleozoic marine environments during the antediluvian period. Personal preliminary research does seem to indicate that the European Upper Triassic reefs sit on Upper Paleozoic sedimentary rocks or directly on Precambrian basement rocks. They do not sit on Middle to Lower Paleozoic rocks. In other words, both ecosystems may have been contemporaneous but geographically separated.
Better Explanatory Power
The model proposed here better explains other data. Some biostratigraphic and paleontological challenges associated with the consensus-view of the Flood, for example, can be mitigated using this model. According to many creationists, the entire North American craton was eroded to an almost flat surface, with continental margins experiencing some of the deepest erosion—perhaps as deep as 1 km, all in just one year (Baumgardner 2018). According to John Baumgardner (2018), this erosion required thousands of tsunamis with wave amplitudes of several hundred feet that pounded the North American craton every few minutes. Baumgardner has conceded that his model is simply a starting point (personal communication from a colleague), but a scenario even remotely similar to this is difficult to reconcile with the paleontological evidence. Given the model proposed here, however, Paleozoic sediments were already in place at the time of the Flood-proper, which means Mesozoic erosion and deposition may have been less erosive and more orderly. Such a scenario seems to be required given the relationship between contemporary Mesozoic dinosaur fossil assemblages. Young earth creationist Matt McLain explains:
For instance, in the Campanian of the western United States, the same dinosaur families are present in Alberta (Dinosaur Park Formation and Oldman Formation), Montana (Judith River Formation and Upper Two Medicine Formation), Utah (Wahweap Formation and Kaiparowits Formation), and New Mexico (Kirtland Formation), including Tyrannosauridae, Hadrosauridae, Ceratopsidae, and Pachycephalosauridae. However, the species present in each of these locations are different. Only a few species are shared between the different states or provinces. For the most part, species are endemic to a particular state or province, even if they are of the same geologic stage and in the same region of the continent. Thus, it appears that the taxa preserved together in these formations represent taxa that lived together in some form before death. I do not think it is likely that these organisms are mixed from different communities because the species are endemic even though the same families are represented across the whole region. Thus, these formations (and I would argue the Maastrichtian above and possibly Santonian below) from western North America represent – as a whole – a particular community type. That is to say, there was an archetypal community in the pre-Flood world that had tyrannosaurids, hadrosaurids, ceratopsids, and pachycephalosaurids as major constituents. There were different actual communities following this formula that are today preserved across North America in the uppermost Cretaceous (unpublished; emphasis mine. See also Wise 2011).
There is also the Cambrian Explosion. The contrast between Cambrian and Precambrian life is conspicuous and striking. According to the consensus creationist view, the virtual absence of metazoan fauna in the Precambrian is due to low burial potential in pre-Flood environments (Snelling 2009, 709). But this solution is tenuous at best. Given the presence of ubiquitous pre-Flood streams and rivers, in conjunction with the existence of large sequences of Neoproterozoic sandstones and conglomerates indicative of energetic sedimentary processes, shouldn’t we expect to find just one fossilized mollusk, or just one fossilized brachiopod? The lack of even a single shelled organism in the Precambrian is better explained as a consequence of special creation in conjunction with an historical curse rather than the Noahic Flood. This solution lends more credibility to the utter disparity that exists between two suites of strata separated by just millimeters of rock.
In a way, this model really just “stretches out” the traditional creationist Flood model from a single year to perhaps as many as 1,600. This “stretching out,” however, allows creationists to apply modern environmental and ecological analogues to the rock and fossil records for the purpose of building high-resolution depositional models. Right now, most Paleozoic and Mesozoic sediments are simply termed “Flood deposits” by many creationists and lack any real environmental or ecological significance.
Depending on where one places the Flood-proper, this model can also be used to build rapid biological diversification models. Such models are impossible if Paleozoic rocks were deposited in just a few months (Brand 2007 agrees). But if these rocks were deposited over 1,600 years, or even 120 years, then rapid diversification is entirely possible. Other areas of creationist research may also benefit from adopting this model.
Conclusion
Thick, geographically extensive, and globally significant assemblages of Cambrian microbialites have been well-documented for over 40 years, but especially over the past ten years. Although only touched on in this paper, vast assemblages of Ordovician microbialite-bearing framework reefs are also globally prevalent and are even more ubiquitous than those in the Cambrian. Other, early Cambrian Archaeocyathid-microbial reefs also exist but were not discussed at all in this paper. Yet each of these microbial-bearing assemblages sit stratigraphically upon the next in multi-km-thick carbonate sequences around the world (figs. 3 and 7). Any attempt to invoke rapid growth and/or allochthonous solutions are fraught with unreasonable, non-scientifically-based assumptions that are unsatisfactory.
That these reefs grew in place is the simplest explanation. This, of course, requires creationists to re-think a Precambrian-Cambrian pre-Flood/Flood boundary. I suggest that most Great Unconformity locations formed during Creation Week, and that vast, epeiric seas existed as original, specially created ecosystems. Early Paleozoic ecosystems thrived in these continental shallow seas but were successively displaced by off craton fauna due to the advent of rapid plate movements. With the onset of the Noahic Flood proper, these communities were finally destroyed. The Flood proper more than likely began during Carboniferous–Permian time or perhaps as late as the Early Mesozoic.
Importantly, this model in no way diminishes catastrophism. Yes, I am proposing that much of the Paleozoic geologic record was deposited over 120 to as many as 1,600 years but this is a far cry from hundreds of millions of years! Even though accepting of some secularist scientific hypotheses this model is, therefore, strictly catastrophist in nature. As such, it in no way diminishes the importance of catastrophist arguments such as: rapid erosion of continental sediments, rapid rates of sediment deposition, large-scale soft-sediment deformation features, flat and geographically extensive contacts between major formations, the nature of the Cambrian Explosion, catastrophic plate tectonics, etc. In many respects, this model actually enhances the creationist position.
Finally, I realize that this paper is not going to be popular within the creationist academic community. As I have said to many of my colleagues, this is not the paper that I wanted to write. It is simply the paper I had to write. As such, I ask for much grace. This is a low resolution, big picture model that obviously has flaws, if not fundamental weaknesses. I realize that many papers will subsequently be written defending the traditional creationist perspective: I welcome them. That is what science is all about. My hope is that as these papers are being written, the stromatolite-growth data would be tackled first, before tearing the model apart elsewhere. I would also ask, since this document is exceptionally long, that future dissenters read this paper through very carefully, perhaps multiple times! I ask this because my writings have already been grossly misunderstood by other creationists who simply failed to read them very carefully.13
Acknowledgements
I want to thank my faithful wife, Beth, for her patience, and for allowing me the time to work on this manuscript. Her insight, wisdom, and editorial support has been invaluable. Thank you to my good friends and colleagues, Matt McLain, John Whitmore, and Andrew Snelling, for their helpful comments, editorial assistance, and cautionary advice, even though they do not agree with my model. There has been much tussling back and forth, but I am thankful for their sincere and humble guidance.
References
Adachi, Natsuko, Jianbo Liu, and Yoich Ezaki. 2013. “Early Ordovician Reefs in South China (Chenjiahe Section, Hubei Province): Deciphering the Early Evolution of Skeletal-Dominated Reefs.” Facies 59, no. 2 (April): 451–466.
Aitken, J. D. 1967. “Classification and Environmental Significance of Cryptalgal Limestones and Dolomites, with Illustrations From the Cambrian and Ordovician of Southwestern Alberta.” Journal of Sedimentary Research 37, no. 4 (December 1): 1163–1178.
Allwood, Abigail C., Malcolm R. Walter, Ian W. Burch, and Balz S. Kamber. 2007. “3.43 Billion-Year-Old Stromatolite Reef from the Pilbara Craton of Western Australia: Ecosystem-Scale Insights to Early Life on Earth.” Precambrian Research 158, nos. 3–4 (5 October): 198–227.
Austin, Steven A., and Kurt P. Wise. 1994. “The Pre-Flood/Flood boundary: As Defined in Grand Canyon, Arizona and Eastern Mojave Desert, California.” In Proceedings of the Third International Conference on Creationism. Edited by R. E Walsh, 37–47. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Baumgardner, John R. 1990. “3-D Finite Element Simulation of the Global Tectonic Changes Accompanying Noah’s Flood.” In Proceedings of the Second International Conference on Creationism. Edited by R. E. Walsh and C. L. Brooks, 35–44. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Baumgardner, John R. 2000. “Distribution of Radioactive sotopes in the Earth.” In Radioisotopes and the Age of the Earth: A Young-Earth Creationist Research Initiative. Edited by Larry Vardiman, Andrew A. Snelling, and Eugene F. Chaffin, 49–94. El Cajon, California: Institute for Creation Research; and St. Joseph, Missouri: Creation Research Society.
Baumgardner, John. 2018. “Understanding How the Flood Sediment Record Was Formed: The Role of Large Tsunamis.” In Proceedings of the Eighth International Conference on Creationism. Edited by J. H. Whitmore, 287–305. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Bechly, Günter. 2020. “The Myth of Precambrian Sponges.” Evolution News, May 12. https://evolutionnews.org/2020/05/the-myth-of-precambrian-sponges/.
Berelson, W. M., F. A. Corsetti, C. Pepe-Ranney, D. E. Hammond, W. Beaumont, and J. R. Spear. 2011. “Hot Spring Siliceous Stromatolites From Yellowstone National Park: Assessing Growth Rate and Laminae Formation.” Geobiology 9, no. 5 (September): 411–424.
Brand, Leonard. 2007. “Wholistic Geology: Geology Before, During, and After the Biblical Flood.” Origins 61 (June 1): 7–34.
Brand, L. R., S. E. Phillips, and A. V. Chadwick. 2012. “Elongated Stromatolite-Like Structures in the Upper Cambrian Hellnmaria Member, Notch Peak Formation, Utah.” Geological Society of America, Abstracts with Programs 44, no. 7: 22.
Burne, Robert V., and Linda S. Moore. 1987. “Microbialites: Organosedimentary Deposits of Benthic Microbial Communities.” Palaios 2, no. 3: 241–254.
Campbell, John A. 1976. “Upper Cambrian Stromatolitic Biostrome, Clinetop Member of the Dotsero Formation, Western Colorado.” Geological Society of America Bulletin 87, no.9 (September): 1331–1335.
Chen, Jitao, Jeong-Hyun Lee, and Jusun Woo. 2014. “Formative Mechanisms, Depositional Processes, and Geological Implications of Furongian (Late Cambrian) Reefs in the North China Platform.” Palaeogeography, Palaeoclimatology, Palaeoecology 414 (15 November): 246–259.
Chen, Zhe, Chuanming Zhou, Xunlai Yuan, and Shuhai Xiao. 2019. “Death March of a Segmented and Trilobate Bilaterian Elucidates Early Animal Evolution.” Nature 573, no. 7774 (19 September): 412–415.
Choh, Suk-Joo, Jongsun Hong, Ning Sun, Sung-Wook Kwon, Tae-Yoon Park, Jusun Woo, Yi Kyun Kwon, Dong-Chan Lee and Dong-Jin Lee. 2013. “Early Ordovician Reefs From the Taebaek Group, Korea: Constituents, Types, and Geological Implications.” Geosciences Journal 17, no. 2 (June): 139–149.
Clarey, Tim. 2018. “Stromatolites: Evidence of Pre-Flood Hydrology.” Acts & Facts 47, no. 5 (April 30)
Corsetti, Frank, A., and Alan J. Kaufman. 2003. “Stratigraphic Investigations of Carbon Isotope Anomalies and Neoproterozoic Ice Ages in Death Valley, California.” GSA Bulletin 115, no. 8 (August): 916–932.
Coulson, Ken P., and Leonard R. Brand. 2016. “Lithistid Sponge-Microbial Reef-Building Communities Construct Laminated, Upper Cambrian (Furongian) ‘Stromatolites.’” Palaios 31, no. 7 (July): 358–370.
Coulson, Ken P., L.R. Brand, and A.V. Chadwick. 2016. “Microbialite Elongation By Means of Coalescence: An Example From the Middle Furongian (Upper Cambrian) of Notch Peak formation of Western Utah.” Facies 62, no. 3 (July). https://doi.org/10.1007/s10347-016-0469-5.
Coulson, Ken P. 2018. “Global Deposits of in situ Upper Cambrian Microbialites: Implications For a Cohesive Model of Origins.” In Proceedings of the Eighth International Conference on Creationism. Edited by J. H. Whitmore, 379– 388. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Coulson, K. P. 2020. Creation Unfolding: A New Perspective on Ex Nihilo, 192. Phaneros Press.
Darroch, Simon A. F., Emily F. Smith, Marc Laflamme, and Douglas H. Erwin. 2018. “Ediacaran Extinction and Cambrian Explosion.” Trends in Ecology and Evolution 33, no. 9 (September): 653–663.
Demicco, Robert V. 1985. “Platform and Off-Platform Carbonates of the Upper Cambrian of Western Maryland, U.S.A.” Sedimentology 32, no. 1 (February): 1–22.
Dickens, Harry, and Andrew A. Snelling. 2008. “Precambrian Geology and the Bible: A Harmony.” Journal of Creation 22, no. 1 (April): 65–72.
Dickens, Harry. 2017. “Colossal Water Flows During Early Creation Week and Early Flood.” Answers Research Journal 10 (September 27): 221–235. https://answersingenesis.org/geology/sedimentation/colossal-water-flows-during-earlycreation-week-and-early-flood/.
Dickens, Harry. 2018. “North American Precambrian Geology—A Proposed Young Earth Biblical Model.” In Proceedings of the Eighth International Conference on Creationism. Edited by J. H. Whitmore, 389–403. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Eagan, K. E., and W. D. Liddell. 1997. “Stromatolite biostromes as bioevent horizons; an example from the Middle Cambrian Ute Formation of the eastern Great Basin.” In Paleontological Events: Stratigraphic, Ecological, and Evolutionary Implications. Edited by Carlton E. Brett, and Gordon C. Baird, 285–308. New York, New York: Columbia University Press.
Faulkner, Danny R. 2016. The Created Cosmos: What the Bible Teaches About Astronomy. Green Forest, Arkansas: New Leaf Publishing Group.
Gebelein, Conrad D. 1969. “Distribution, Morphology, and Accretion Rate of Recent Subtidal Algal Stromatolites, Bermuda.” Journal of Sedimentary Research 39, no. 1 (March 1): 49–69.
Gould, Stephen Jay. 2002. The Structure of Evolutionary Theory. Cambridge, Massachusetts: Belknap Harvard.
Hagadorn, James W., and Ben Waggoner. 2000. “Ediacaran Fossils From the Southwestern Great Basin, United States.” Journal of Paleontology 74, no. 2 (March): 349–359. https://bioone.org/journals/journal-of-paleontology/volume-74/issue-2
Hong, Jongsun, Seong-Hyeon Cho, Suk-Joo Choh, Jusun Woo, and Dong-Jin Lee. 2012. “Middle Cambrian Siliceous Sponge-Calcimicrobe Buildups (Daegi Formation, Korea): Metazoan Buildup Constituents In the Aftermath Of the Early Cambrian Extinction Event.” Sedimentary Geology 253–254 (1 May): 47–57.
Horstemeyer, Mark, and John R. Baumgardner. 2003. “What Initiated the Flood Cataclysm?” In Proceedings of the Fifth International Conference on Creationism. Edited by R. L. Ivey Jr., 155–163. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Hintze, Lehi F., Michael E. Taylor, and James F. Miller. 1988. “Upper Cambrian–Lower Ordovician Notch Peak Formation in Western Utah.” United States Geological Survey Professional Paper 1393: 1–29.
Hintze, Lehi F., and Fitzhugh D. Davis. 2003. “Geology of Millard County, Utah.” Utah Geological Survey Bulletin 133: 305 p.
Hughes, Nigel C., and Dan L. Cooper. 1999. “Paleobiologic and Taphonomic Aspects Of the ‘Granulosa’ Trilobite Cluster, Kope Formation (Upper Ordovician, Cincinnati Region).” Journal of Paleontology 73, no. 2 (20 May): 306–319.
Johns, Ronald A., Benjamin F. Dattilo, and Ben Spincer. 2007. “Neotype and Redescription of the Upper Cambrian Anthaspidellid Sponge, Wilbernicyathus donegani Wilson, 1950.” Journal of Paleontology 81, no. 3 (May): 435–444.
Johns, Warren H. 2016. “Scriptural Geology, Then and Now.” Answers Research Journal 9 (November 30): 317–337. https://answersingenesis.org/age-of-the-earth/scripturalgeology-then-and-now/
Johnson, James J. S. 2011. “Biblical Devastation in the Wake of a ‘Tranquil Flood.’” Acts and Facts, 40, no. 9: 8–10.
Karlstrom, K. E., M. T. Mohr, M. D. Schmitz, F. A. Sundberg, S. M. Rowland, R. Blakey, J. R. Foster, L. J. Crossey, C. M. Dehler, and J. W. Hagadorn. 2020. “Redefining the Tonto Group of Grand Canyon and Recalibrating the Cambrian Time Scale.” Geology 48, no. 5 (1 May): 425–430.
Kennard, John M., and Noel P. James. 1986. “Thrombolites and Stromatolites: Two Distinct Types of Microbial Structures.” Palaios 1, no. 5 (October): 492–503.
Knight, I., Karem Azmy, William Douglas Boyce, and Denis Lavoie. 2008. “Tremadocian Carbonate Rocks of the Lower St. George Group, Port au Port Peninsula, Western Newfoundland: Lithostratigraphic Setting of Diagenetic, Isotopic, and Geochemical Studies.” Current Research. Report 08-1. Geological Survey, 1-43. St. John’s, Newfoundland: Newfoundland and Labrador Department of Natural Resources.
Kruse, Peter D., and Joachim R. Reitner. 2014. “Northern Australian Microbial-Metazoan Reefs After the Mid- Cambrian Mass Extinction.” Memoirs of the Association of Australasian Paleontologists 45 (April): 31–53.
Lee, Hyun Suk, and Sung Kwun Chough. 2011. “Depositional Processes of the Zhushadong and Mantou Formations (Early to Middle Cambrian), Shandong Province, China: Roles of Archipelago and Mixed Carbonate–Siliciclastic Sedimentation on Cycle Genesis During Initial Flooding of the North China Platform.” Sedimentology 58, no. 6 (October): 1530–1572.
Lee, Jeong-Hyun, Jitao Chen, and Sung Kwun Chough. 2015. “The Middle-Late Cambrian Reef Transition and Related Geological Events: A Review and a New View.” Earth- Science Reviews 145 (June): 66–84.
Lee, Jeong-Hyun, Jitao Chen, and S. K. Chough. 2010. “Paleoenvironmental Implications Of an Extensive Maceriate Microbialite Bed In the Furongian Chaomidian Formation, Shandong Province, China.” Palaeogeography, Palaeoclimatology, Palaeoecology 297, nos. 3–4 (20 November): 621–632.
Li, Qijian, Yue Li, Jianpo Wang, and Wolfgang Kiessling. 2015. “Early Ordovician Lithistid Sponge-Calathium Reefs on the Yangtze Platform and Their Paleoceanographic Implications.” Palaeogeography, Palaeoclimatology, Palaeoecology 425 (1 May): 84–96.
Logan, Brian W. 1961. “Cryptozoon and Associate Stromatolites from the Recent, Shark Bay, Western Australia.” Journal of Geology 69, no. 5 (September): 517–533.
Miller, James F., Kevin R. Evans, James D. Loch, Raymond L. Ethington, James H. Stitt, Lars Holmer, and Leonid E. Popov. 2003. “Stratigraphy of the Sauk III Interval (Cambrian–Ordovician) In the Ibex Area, Western Millard County, Utah and Central Texas.” Brigham Young University Geological Studies 47: 23–141.
Miller, James F., Kevin R. Evans, and Benjamin F. Dattilo. 2012. “The Great American Carbonate Bank in the Miogeocline of Western Central Utah: Tectonic Influences on Sedimentation.” In The Great American Carbonate Bank: The Geology and Economic Resources of the Cambrian– Ordovician Sauk Megasequence of Laurentia. Edited by James Derby, Richard Fritz, Susan Longacre, William Morgan, and Charles Sternbach. American Association of Geologists memoir 98: 769–854.
Miller, James F., James D. Loch, and John F. Taylor. 2012. “Biostratigraphy of Cambrian and Lower Ordovician Strata in the Llano Uplift, Central Texas.” In The Great American Carbonate Bank: The Geology and Economic Resources of the Cambrian-Ordovician Sauk Megasequence of Laurentia. Edited by James Derby, Richard Fritz, Susan Longacre, William Morgan, and Charles Sternbach. American Association of Geologists memoir 98: 187–202.
Mortenson, Terry. 1998. “Andrew Ure (1778–1857).” https://answersingenesis.org/creation-scientists/profiles/andrewure/.
Peters, Shanan E., and Robert R. Gaines. 2012. “Formation Of the ‘Great Unconformity’ As a Trigger For the Cambrian Explosion.” Nature 484, no. 7394 (19 April): 363–366.
Peters, Shanan E., Jon M. Husson, and Julia Wilcots. 2017. “The Rise and Fall of Stromatolites in Shallow Marine Environments.” Geology 45, no. 6 (June): 487–490.
Petroff, Alexander P., Min Sub Sim, Andrey Maslov, Mikhail Krupenin, Daniel H. Rothman and Tanja Bosak. 2010. “Biophysical Basis For the Geometry of Conical Stromatolites.” Proceedings of the National Academy of Science 107, no. 22 (June 1): 9956–9961.
Playford, P. E. 1980. “Environmental controls on the morphology of modern microbialites at Hamelin Pool, Western Australia.” Geological Survey of Western Australia. Annual Report for 1979: 73–77.
Powell, D. Keith. 1959. “The Geology of the Southern House Range Millard County, Utah.” Brigham Young University Research Studies 6, no. 1, Brigham Young University. M.Sc. thesis.
Purdom, Georgia, and Andrew A. Snelling. 2013. “Survey of Microbial Composition and Mechanisms of Living Stromatolites Of the Bahamas and Australia: Developing Criteria to Determine the Biogenicity of Fossil Stromatolites.” In Proceedings of the Seventh International Conference on Creationism. Edited by M. Horstemeyer. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Reid, R. P, P. T. Visscher, A. W. Decho, J. F Stolz, B. M Bebout, C. Dupraz, I. G Macintyre et al. 2000. “The Role of Microbes in Accretion, Lamination and Early Lithification of Modern Marine Stromatolites.” Nature 406, no. 6799 (31 August): 989–992.
Robbins, L. L., Y. Tao, and C. A. Evans. 1997. “Temporal and Spatial Distribution of Whitings on Great Bahama Bank and a New Lime Mud Budget.” Geology 25, no. 10 (October): 947–950.
Robinson, Steven J. 1996. “Can Flood Geology Explain the Fossil Record?” Creation Ex Nihilo Technical Journal 10, no. 1 (April): 32–69.
Saltzman, Matthew R., Clinton A. Cowan, Anthony C. Runkel, Bruce Runnegar, Michael C. Stewart, and Allison R. Palmer. 2004. “The Late Cambrian SPICE (δ13C) Event and the Sauk II–Sauk III Regression: New Evidence From Laurentian Basins in Utah, Iowa, and Newfoundland.” Journal of Sedimentary Research 74, no. 3 (May): 366–377.
Shapiro, Russell S., and Stanley M. Awramik. 2000. “Microbialite Morphostratigraphy as a Tool for Correlating Late Cambrian–Early Ordovician Sequences.” The Journal of Geology 108, no. 2 (March): 171–180.
Sigler, Roger and Van Wingerden. 1998. “Submarine Flow and Slide Deposits in the Kingston Peak Formation, Kingston Range, Mojave Desert, California: Evidence for Catastrophic Initiation of Noah’s Flood.” In The Proceedings of the International Conference on Creationism. Vol. 4.
Snelling, Andrew A. 2008. “Thirty Miles of Dirt in a Day.” Answers Magazine 3, no. 4 (October–December): 29–30.
Snelling, Andrew A. 2009. Earth’s Catastrophic Past: Geology, Creation and the Flood. Part II. Dallas, Texas: Institute of Creation Research.
Srinivasan, Krishnan, and Kenneth R. Walker. 1993a. “Sequence Stratigraphy Of an Intrashelf Basin Carbonate Ramp to Rimmed Platform Transition: Maryville Limestone (Middle Cambrian), Southern Appalachians.” GSA Bulletin 105, no. 7 (July 1): 883–896.
Stanley, Steven M., and John A Luczaj. 2015. Earth System History. 4th ed. New York, New York: W. H. Freeman and Company.
Taylor, John F., John E. Repetski, James D. Loch, and Stephen A. Leslie. 2012. “Biostratigraphy and Chronostratigraphy of the Cambrian–Ordovician Great American Carbonate Bank.” In The Great American Carbonate Bank: The Geology and Economic Resources of the Cambrian- Ordovician Sauk Megasequence of Laurentia. Edited by James Derby, Richard Fritz, Susan Longacre, William Morgan, and Charles Sternbach. American Association of Geologists Memoir 98: 187–202.
Tóth, L. Fejes. 1948. “On the Densest Packing of Circles in a Convex Domain.” Norske Vid. Selsk. Fordhl., Trondheim 21: 68–76.
Whitcomb, John C., and Henry M. Morris. 1961. The Genesis Flood: The Biblical Record and its Scientific Implications. Phillipsburg, New Jersey: P&R Publishing.
Whitmore, John, H. 2009. “Modern and Ancient Reefs.” In Rock Solid Answers: The Biblical Truth Behind 14 Geologic Questions. Edited by Mike Oard and John K. Reed, 149– 166. Green Forest, Arkansas: Master Books.
Whitmore, John, H., and Paul Garner. 2008. “Using Suites of Criteria to Recognize Pre-Flood, Flood, and Post-Flood Strata in the Rock Record with Application to Wyoming (USA).” In Proceedings of the Sixth International Conference on Creationism. Edited by Andrew A. Snelling, 425–448. Pittsburgh, Pennsylvania: Creation Science Fellowship; and Dallas, Texas: Institute for Creation Research.
Wise, Kurt P. 2011. “Noah’s World—Same Time, Different Place.” Answers Magazine 6, no. 4 (September 26), 56–63.
Wise, Kurt P., and Andrew A. Snelling. 2005. “A Note On the pre-Flood/Flood Boundary in the Grand Canyon.” Origins 58 (June 1): 7–29.
Woo, J., and S. K. Chough. 2010. “Growth Patterns Of the Cambrian Microbialite: Phototropism and Speciation of Epiphyton.” Sedimentary Geology 229, nos. 1–2 (15 July): 1–8.
Wood, Rachel. 1999. Reef Evolution Oxford, United Kingdom: Oxford University Press, Oxford.
Woods, A. D. 2009. “Anatomy of An Anachronistic Carbonate Platform: Lower Triassic Carbonates of the Southwestern United States.” Australian Journal of Earth Sciences 56, no. 6 (13 July): 825–839.
Yonkee, W. A., C. D. Dehler, P. K. Link, E. A. Balgord, J. A. Keeley, D. S. Hayes, M. L. Wells, C. M. Fanning, and S. M. Johnston. 2014. “Tectono-Stratigraphic Framework of Neoproterozoic to Cambrian Strata, West-Central U.S.: Protracted Rifting, Glaciation, and Evolution of the North American Cordilleran Margin.” Earth-Science Reviews 136 (September): 59–95.















