The views expressed in this paper are those of the writer(s) and are not necessarily those of the ARJ Editor or Answers in Genesis.
Abstract
Many evolutionists claim that dinosaurs evolved into birds. Since this claimed evolutionary transition necessitates the concept that a land-based creature developed the ability to fly, aeronautical considerations such as center of gravity should be considered. Birds have an anteriorly placed center of gravity, near the chest, enabling pitch stability in flight, meaning that a bird’s nose up or nose down pitch angle is stable and controllable. Theropod dinosaurs have a posteriorly placed center of gravity, close to the hip joint, which runs counter to pitch stability in flight. This author conducted a literature review of evolutionary research regarding claims of dinosaur-to-bird evolutionary, anterior shift in center of gravity. Review of the evolutionary literature at the time of this writing reveals reliance on several hypotheses regarding supposed morphological changes to shift the center of gravity, and questionable labeling of birds as dinosaurs as evidence that the center of gravity did indeed shift. The goals of this paper are to form a hypothesis that birds and theropod dinosaurs have always had distinct differences in center of gravity, make predictions from that hypothesis, and recommend topics for further research.
Keywords: dinosaurs, birds, flight, stability, evolution, mass, weight, center of gravity, pitch
Introduction
To clarify terminology up front, this author uses the 1881 definition of “theropod” rather than the 1986 definition. Haynes (2022) explains the difference:
In 1881, Othniel Charles Marsh created the Theropoda suborder (now clade), grouping all known dinosaurs from the Triassic and the carnivorous dinosaurs from the Jurassic and Cretaceous. Jacques Gauthier described theropods in 1986 via cladistics (an evolutionary method that infers ancestry) as a group of birds and all saurischians (dinosaurs).
Many evolutionists claim that dinosaurs evolved into birds, necessitating significant morphological changes. Such changes include the development and aerodynamic arrangement of feathers, reengineering of forelimbs, pectoral girdle, hips, tail, respiratory system, and much more. Creationists are conducting ongoing research into the significant problems and limitations of these irreducibly complex biological changes. The complexities of these hypothesized biological changes are further compounded when combined with aeronautical considerations, such as the placement of center of gravity (CoG) in birds versus theropod dinosaurs. Birds have an anteriorly placed (that is, toward the forward end of the body) CoG near the chest, while theropod dinosaurs have a posteriorly placed (that is, toward the aft end of the body) CoG near the hip joint. This author hypothesizes that the CoG distinction between birds (not necessarily flightless birds) and theropod dinosaurs has always been distinct, and offers predictions and recommendations for future research. This hypothesis provides an aeronautical engineering consideration to a subject that is otherwise typically dominated by fields of study such as paleontology, biomechanics, and evolutionary biology. While the topic of dinosaur-to-bird evolution necessitates those typical fields of study, aeronautical considerations such as CoG provide perspectives that can add value to ongoing creationist research regarding birds and dinosaurs.
Flight operations personnel and aeronautical engineers are usually well trained to understand the critical importance of CoG placement with respect to aerodynamic forces and moments. Nelson (1998) derives an equation for the total pitching moment of an airplane called the stick fixed neutral point. The stick fixed neutral point is a function of pitching moment contributions from the aircraft wing, fuselage, and tail contributions. The neutral point tends to be close to the wings on a typical aircraft that has a horizontal stabilizer in the rear (that is, the aircraft has a tail). Oftentimes, scientists and engineers will compare the CoG location with that of the center of lift of the wing for simplicity, since the neutral point tends to be close to the wings, but note that pitch stability ultimately depends on the location of the CoG relative to the neutral point. Nelson (1998) explains how the derivative (slope) of pitching moment coefficient (Cm) versus angle of attack (α) must be less than zero (negative) for pitch stability (dCm/dα < 0). Such longitudinal stability requires the CoG to be placed at or forward of the neutral point. If the CoG is aft of the neutral point, then this results in a positive derivative of Cm versus α due to destabilizing moment arms, leading to unstable pitch. See fig. 1 for a visual summary of dCm/dα. See fig. 2 for a visual summary of CoG location relative to the neutral point on an aircraft.
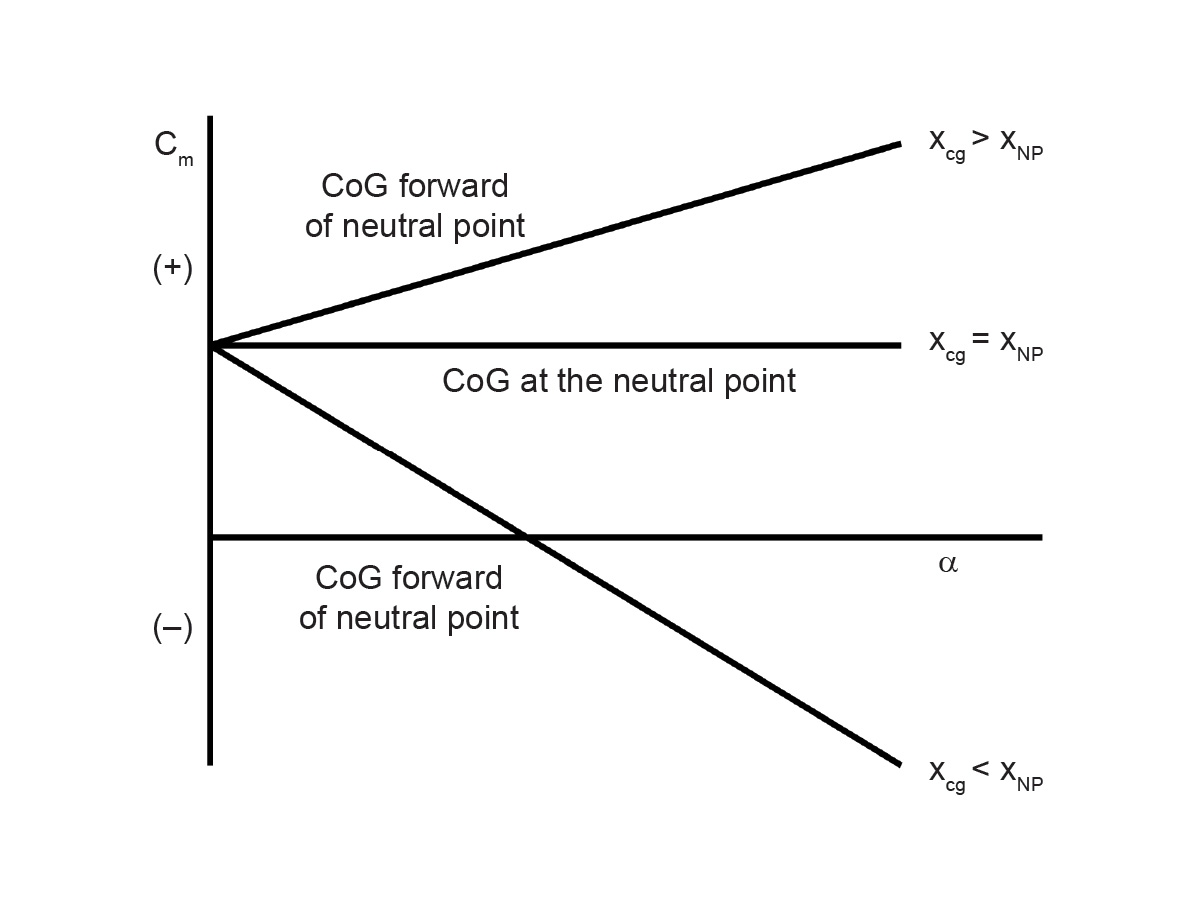
Fig. 1. The influence of CoG position on longitudinal static stability” Adapted from Nelson (1998, 56), figure 2.15.
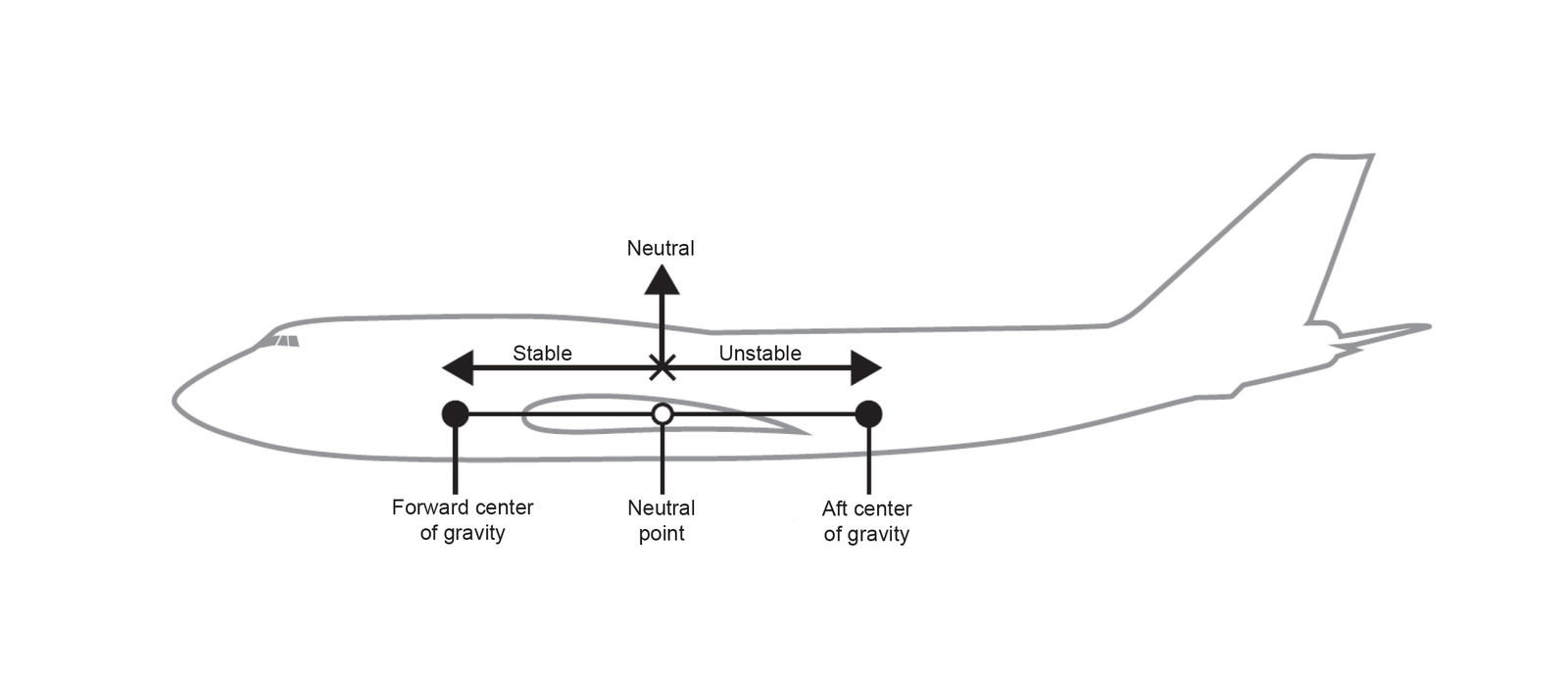
Fig. 2. “The neutral point compared to stable and unstable CoG locations on an aircraft” Adapted from Tecuceanu (2023, 249), figure 2.
Flight operations personnel are aware of CoG limitations, which is why they must comply with aircraft model-specific weight and balance specifications. Aeronautical engineers must consider weight and balance not only in the design of new aircraft, but for aircraft modifications to ensure that such modifications comply with weight and balance-related airworthiness requirements. Failure to comply with weight and balance specifications and airworthiness requirements could result in unstable flight characteristics, loss of control, and catastrophic failure.
An aircraft can have all the necessary features and controls for flight—engines for thrust, wings for lift, flight controls and control surfaces, avionics, and so forth. Yet, if the CoG is shifted too far aft, then the aircraft is likely to enter an unstable pitch right after takeoff, making the aircraft prone to uncontrollable nose-up tendencies, increased risk of aerodynamic stall (this occurs when the angle of attack exceeds the critical angle), and loss of control. The same aircraft stability physics applies to flying creatures—pitching moments must be properly balanced for pitch stability in flight.
Many of the supposed dinosaur-to-bird transitional forms are artistically reconstructed as dinosaurs with feathered wings. Creationist research on the topic of supposed dinosaurs with feathers is ongoing, such as the works of Sanders (2025), and is not the focus or purpose of this paper. This paper focuses on the necessity of the anterior CoG placement for flight and how this pertains to the debate regarding dinosaur-to-bird evolution. The literature review and analysis demonstrates that evolutionists acknowledge the necessity of the CoG closely aligning with the center of lift to enable stable flight. Evolutionary researchers offer several explanations as to how the CoG placement evolved over time, and this paper will critique those explanations. Evolutionary researchers also point to the fossil record and utilize cladistics to support their claim that CoG evolution occurred. This paper will examine some of those claims, but cladistics analysis is not the focus of this paper. Lastly, this author makes predictions and provides recommendations for future research pertaining to this topic.
Methodology
This paper examines secular published research related to dinosaur-to-bird evolutionary claims on the topic of CoG. Not every single paper in existence that mentions CoG is cited and analyzed, but only those papers that offer the most research and analysis regarding the importance of CoG placement for flight and bipedalism, as well as the papers that hypothesize mechanisms on how the CoG location shifted in dinosaur-to-bird evolution. Note that this paper generally does not question calculated CoG locations for fossilized, extinct creatures, except when such calculations might be tainted by evolutionary assumptions. This paper critiques some evolutionary claims of fossil evidence that supposedly aligns with evolutionary shift in CoG, but it is not meant to be an exhaustive cladistical analysis. This literature review is used to support the hypothesis of this author that God created birds and theropod dinosaurs with distinct CoG locations: birds with an anterior CoG, and theropod dinosaurs with a posterior CoG. This paper uses that hypothesis to make several predictions, and this author offers recommendations for future creationist research to augment the dinosaur-to-bird paleontological and biological debate with this important aeronautical consideration.
CoG Importance Acknowledged by Evolutionists
The need for the CoG to be close (positioned slightly forward) to the center of lift is well established not just for aircraft, but also for flying creatures. Within the context of Pteranodon flight, Bramwell and Whitfield (1974, 541) state the following:
for in steady flight the centre of lift and the centre of gravity must coincide. If the centre of gravity lies in front of the centre of lift, the animal will nose dive; if it is behind the centre of lift it will stall.
Likewise, Jones et al. (2000, 716) note the following about birds:
Modern birds have markedly foreshortened tails and their body mass is centred anteriorly, near the wings. To provide stability during powered flight, the avian centre of mass is far from the pelvis . . .
Moreover, through observations of living birds and reasonable estimates based on dinosaur fossils, evolutionary scientists acknowledge that the CoG for birds is positioned anteriorly, near their wings (i.e. center of lift), while the CoG for theropod dinosaurs is closer to their hips. For example, Gatesy (1990, 183) states the following regarding CoG/center of mass (CoM):
birds have a center of mass located well in front of the hip joint. Birds maintain a relatively horizontal femur to locate the feet under the center of mass . . . Unlike that of birds, the tail of primitive theropods was a substantial post acetabular fraction of total body weight and would have located the center of mass close to the hip joint.
The term “primitive” in the latter quote is based on an evolutionary assumption that there is such a thing as a primitive dinosaur or primitive bird. Nevertheless, Gatesy correctly acknowledges the CoG/CoM placement near the hip joint for theropod dinosaurs due to their substantial tail mass.
Degernes and Feduccia (2001, 10) also acknowledge an anterior CoG for birds:
Birds are unique among vertebrates in having extraordinarily large flight muscles located on the keeled sternum with a center of gravity ventral to the wings.
Fig. 3 illustrates the difference in estimated CoG location between an extinct bird (Archaeopteryx) and a theropod dinosaur (Sinosauropteryx). Note that many evolutionists may claim that Archaeopteryx is a dinosaur, but there are reasonable grounds to categorize Archaeopteryx as a bird per research conducted by Haynes (2022, 2023). Nevertheless, the main point of this illustration is to contrast the differences in CoG location between birds and theropod dinosaurs.
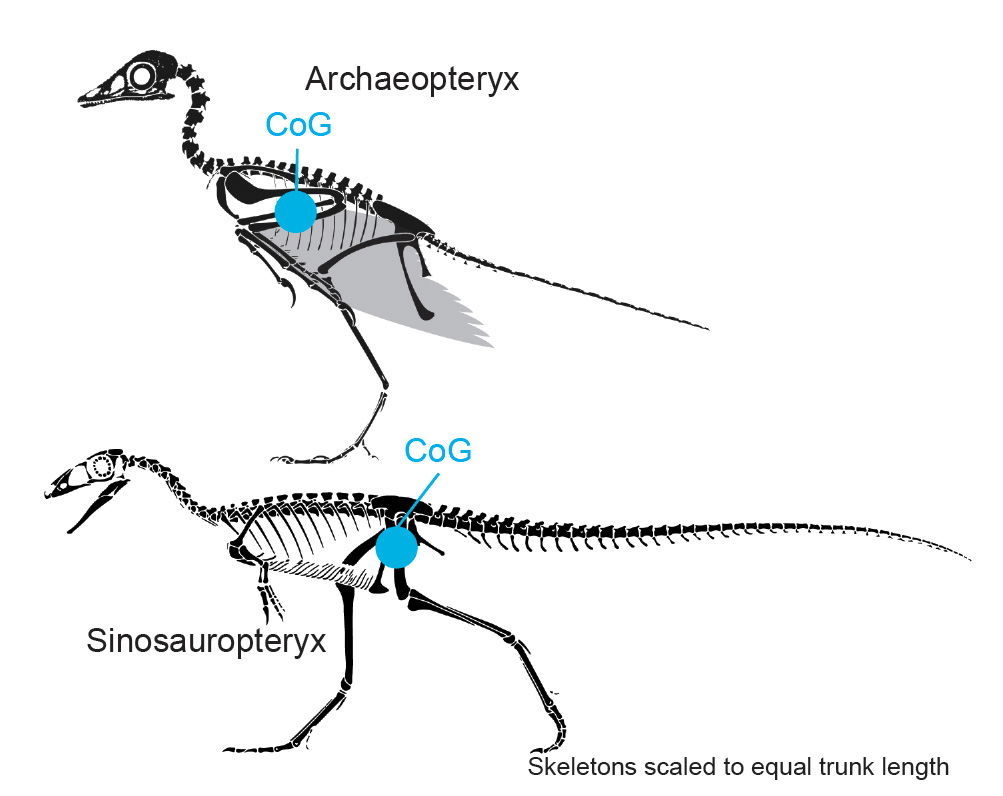
Fig. 3. Estimated CoG location comparison between an extinct bird Archaeopteryx and a dinosaur Sinosauropteryx; skeletal drawing adapted from Scott Hartman (2020) reconstruction. https://www.skeletaldrawing.com/theropods/sinosauropteryx.
Evolutionary Claims Regarding Anterior Shift in CoG
Given the anterior CoG placement in birds contrasted with posterior CoG placement in theropod dinosaurs, evolutionary researchers attempt to theorize how the CoG shifted forward in supposed dinosaur-to-bird evolution. Some evolutionists theorize that dinosaur tail reduction caused the anterior shift in CoG (that is, a shorter tail would decrease the posterior mass, shifting the CoG forward). Carrano (1998, 464) states:
The horizontal posture of avian femora has been linked to an anterior shift in the center of mass associated with tail reduction.
Farlow et al. (2000, 644) similarly claim:
Theropods’ more closely related to birds underwent a reduction in tail size. The above-noted progressive reduction in the number of caudal vertebrae was accompanied by a reduction in tail diameter, particularly distally. These changes likely affected the location of the body’s center of mass, shifting it forward relative to the hip joint and forcing the limbs to reorient to position the feet further forward.
Yet, this supposed evolutionary change (hypothesized, but not directly observed) would have a significant impact on locomotion, as Farlow et al. (2000, 464) explain:
Changes in the size and shape of the tail had additional effects on locomotion. The tail’s length and mass affected the way ‘theropods’ balanced. As obligate bipeds, ‘theropods’ had to stand with the body’s center of mass over the feet to maintain equilibrium. A nose-down pitch caused by the body in front of the hip joint was counterbalanced, at least in part, by the nose-up rotation imparted by the tail.
Hutchinson (2009, 426) notes the CoG difference between hip-driven locomotion for dinosaurs versus the knee-driven locomotion for birds:
To some degree, the hip-driven mechanism seems linked to a more caudally positioned centre of mass of the body, whereas a more cranially positioned centre of mass may be correlated with a more knee-driven mechanism. The evolutionary transition between these mechanisms is illuminated by tail reduction and pectoral limb expansion (thus a presumed centre of mass shift) from basal theropods to birds.
Yet, if the CoG shifted in evolving dinosaurs via tail reduction, such a change would be limited by irreducible complex functionality. Theropod dinosaurs needed their tails for balance, with their CoG located at the hip joint, so any such dinosaur in the evolutionary process of transitioning to an anterior CoG would struggle to walk without numerous other morphological changes that would have to evolve simultaneously, including limb reorientation for knee-driven locomotion. Paul (2016, 29) notes the significant difference in femur orientation and locomotion between dinosaurs and birds:
In bipedal dinosaurs . . . the femur had to slope strongly forward to place the feet beneath the center of gravity. This arrangement is taken to an extreme in short-tailed birds, whose femur is nearly horizontal when they are walking in order to place the knees and feet far enough forward; in running, the femur of birds swings more strongly backward.
Therefore, dinosaurs would need to simultaneously evolve knee-driven locomotion to maintain stability with an anterior CoG. Yet, evolutionary literature and media on the topic of dinosaur-to-bird evolution is proliferated with artwork depicting dinosaurs with feathers while maintaining their dinosaur limb orientation. See fig. 4 as an example.
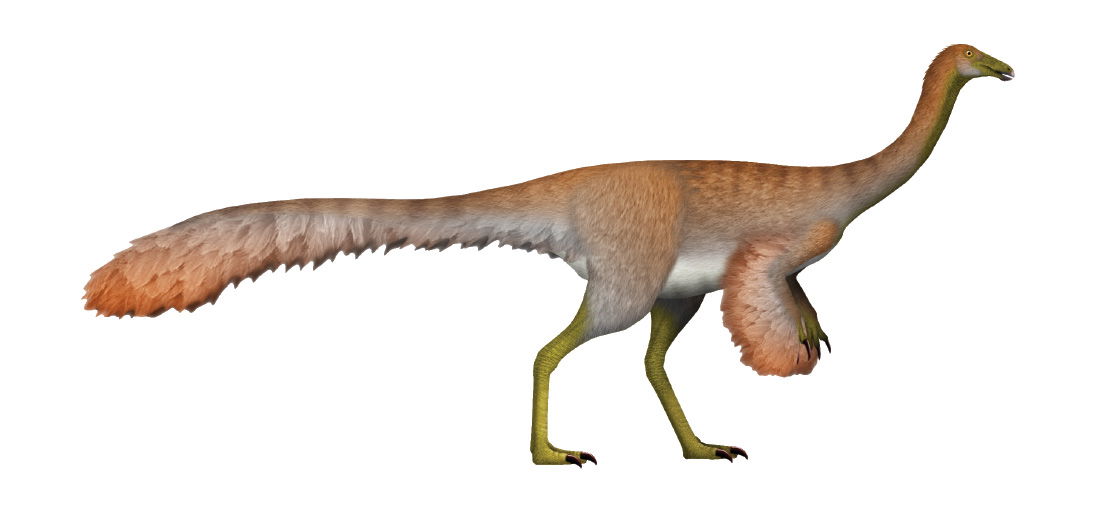
Fig. 4. Example of typical feathered dinosaur artwork. UnexpectedDinoLesson. “Ornithomimus was a swift bipedal theropod which superficially resembled an ostrich. Several specimens have been found preserving evidence of feathers, and it was equipped with a small toothless beak that may indicate an omnivorous diet. Ornithomimus is estimated at about 4 m in length and 170 kg in weight, and is characterized by feet with three weight-bearing toes, long slender arms, and a long neck with a birdlike skull. They had very long limbs, hollow bones, and large brains and eyes.” https://commons.wikimedia.org/wiki/File:Ornithomimus_UDL.png. CC BY-SA 4.0.
Hypothetical dinosaurs illustrated with feathers such as these that would maintain their original morphology (other than the feathers) still have a posterior CoG. With feathers depicted on the forelimbs, the CoG would be far aft of the wing lift. Applying the same physics as that of Nelson (1998) for an airplane, a creature with a CoG aft of the neutral point would exhibit a positive derivative of Cm versus α (dCm/dα > 0) due to destabilizing moment arms. Since the neutral point tends to be close to the center of lift, the creature depicted in fig. 4 would have unstable pitch. Any attempted flight or lift-generating motion (like wing flapping or gliding) would cause the creature to uncontrollably pitch upward and enter an aerodynamic stall.
Some evolutionists have attempted to explain how these morphological changes might have occurred rapidly rather than gradually. Rashid et al. (2014, 25) theorized that mutations in mice that resulted in tail reduction may be relevant to dinosaurs:
To generate a list of candidate genes that may have been modulated in the transition to short-tailed birds, we analyzed a comprehensive set of mouse mutants . . . An open question is whether the relatively sudden appearance of short-tailed birds in the fossil record could be accounted for, at least in part, by the pleiotropic effects generated by a relatively small number of mutational events . . . Interestingly, a prevalent pleiotropic effect of mutations that cause fused caudal vertebral bodies (as in the pygostyles of birds) is tail truncation.
In general, it’s conceivable to see how a mutation can cause tail shortening due to a loss or alteration of genetic information. However, tail shortening is likely not the only change required to turn a dinosaur tail into a bird tail. Bird tails have feathers, and ongoing creationist research, such as the work of Sanders (2025) calls into question many of the feathered dinosaur claims. Ongoing and future research may continue to refute claims of dinosaurs having feathers. If this turns out to be the case, then the evolution of a dinosaur tail into a bird tail must also explain the origin of the feathers. While mutations might cause a tail to shorten, mutations would be unable to account for the evolution of feathers where feathers did not previously exist. First, such a change would require the addition of new, useful genetic information to generate new, phenotypical functionality. Despite evolutionary claims to the contrary, Tan (2022) demonstrated how mutations, gene duplication, and natural selection do not generate genetic diversity. Moreover, Cserhati (2023) demonstrated insurmountable genetic complexities with the claim that scales evolved into feathers due to keratin-related gene similarities. Therefore, tail shortening mutations may be insufficient to demonstrate how a dinosaur tail can evolve into a bird tail, failing to account for the additional genetic/structural changes required.
Compounding the problems further for the evolutionary model, Allen et al. (2013, 105) found that supposed tail reduction would have had a minimal impact on shifting the CoG/CoM anteriorly:
Considering that the tail represents the majority of body mass caudal to the hip, reduction or cranial concentration of tail mass, or both, would be expected to bias the whole-body CoM position strongly cranially. However, our results indicate that the effects of tail reduction were not significant in comparison to concurrent changes to the limbs (especially pectoral) and, to a lesser extent, the head and neck. Therefore, we infer that adding mass to the front of theropod bodies was more influential for CoM evolution than was removing it from the back.
Simply put, if adding mass to the front matters more than removing mass from the back, then tail reduction alone could not have driven the required CoG shift for flight. They even acknowledge the struggle to explain whether purported changes in CoG and body shape came before, during, or after the origin of flight:
Until more robust phylogenetic and aerodynamic assessments for early maniraptoriforms are made, it is impossible to assess conclusively whether our predictions of CoM and body shape change preceded, coincided with or followed the origin of flight. (Allen et al. 2013, 106)
For each of the three ordering possibilities presented by Allen et al. (2013), evolutionists struggle to demonstrate, via observational science, the evolution of every other bird-like feature to include (but not limited to) the following: feathers and their aerodynamic arrangement, forelimb changes to support range of motion for wing flapping, pectoral girdle changes to support flight muscles, respiratory system changes, brain changes to enable spatial awareness, and much more. If evolutionists choose to believe that anterior shift in CoG and body shape changes coincided with the origin of flight, then they must demonstrate how the aforementioned bird-like features also evolved simultaneously, let alone the challenge of demonstrating such evolution gradually and sequentially. If evolutionists choose to believe that CoG and body shape changes followed the origin of flight, then they would have to somehow demonstrate flight with a posterior CoG relative to the neutral point, which contradicts proven and demonstrated aerodynamic physics on the matter such as that of Nelson (1998). If evolutionists choose to believe that an anterior shift in CoG preceded the origin of flight, then they must demonstrate how dinosaurs simultaneously evolved other body shape characteristics to maintain ground locomotion via knee-walking. Some evolutionists have attempted to demonstrate feasibility of the latter option via weighted chicken experiments.
Several researchers experimented with limb posture in chickens to explain supposed evolutionary limb reorientation. These experiments involved adding weights to chicken tails to see if their femoral posture would change to more vertical (like that of dinosaurs) than horizontal. This is essentially attempting to make a bird walk like a dinosaur, so it’s in the opposite direction of the claimed evolutionary change, but the researchers rationalize that demonstrating the feasibility of this change supports the feasibility of an evolutionary change in femur bone in the other direction: from somewhat vertical to horizontal. Moreover, the researchers claim that the orientation of the femur bones impact CoG, and that changes in femoral posture could help explain its evolutionary shift.
After obtaining results in which adding weights caused a more horizontal change (the opposite of what they expected) to the femur, Carrano and Biewener (1999, 248) concluded the following:
the specific postural changes that were induced here do not reflect those hypothesized to have occurred during early bird evolution. Considering the substantial morphological differences between birds and non-avian dinosaurs, this advocates caution in using information derived from modern birds to interpret all aspects of non-avian dinosaur locomotion. Instead, these results emphasize the highly significant degree to which birds have altered limb posture, kinematics, and morphology from that of their terrestrial theropod ancestors.
However, the weighted chicken experiment was revived with Grossi et al. (2014, 1) after making some adjustments to their experiment compared to that of Carrano:
Here we show that, by experimentally manipulating the location of the centre of mass in living birds, it is possible to recreate limb posture and kinematics inferred for extinct bipedal dinosaurs. Chickens raised wearing artificial tails, and consequently with more posteriorly located centre of mass, showed a more vertical orientation of the femur during standing and increased femoral displacement during locomotion . . . Our results support the hypothesis that gradual changes in the location of the centre of mass resulted in more crouched hindlimb postures and a shift from hip-driven to knee-driven limb movements through theropod evolution.
Grossi and colleagues (2014, 5) go on to note a caveat to their findings:
One caveat, however, is that our approach uses tail reduction as the mechanism for CoM displacement despite it has been recently shown that the evolutionary change in CoM position was driven instead by forelimb enlargement [Allen et al. 2013]. Nonetheless, this does not mean that tail reduction had no effect on CoM displacement, but that it was not the most important factor. Ideally we would have increased tail mass and reduced pectoral limb mass but, unfortunately, this is not experimentally feasible.
They go on to conclude:
Thus, we expect that careful phenotypic manipulation of extant birds can open new avenues of experimental investigation into unexplored facets of dinosaur locomotor mechanics and energetics, providing a more nuanced understanding of the relationship between form and function in dinosaur evolution. (Grossi et al. 2014, 5)
While Grossi and colleagues were able to successfully, at least to some extent, make a chicken walk like a dinosaur, such a demonstration accomplished very little in terms of demonstrating the feasibility of dinosaurs evolving from hip-walking to knee-walking. There is no reasonable expectation for such phenotype manipulation to result in any kind of genetic manipulation and genetic inheritance in the offspring, because claims of the sort would be akin to Lamarckian evolution. As noted by Lisle (2018), both creationists and evolutionists generally reject Lamarckian evolution since it contradicts the rules of heredity. Grossi and colleagues did not demonstrate heredity of the altered posture. Moreover, the human-manipulated variables don’t necessarily align with the factors that supposedly drove CoG shift.
While evolutionary researchers struggle to demonstrate the mechanisms for a supposed anterior shift in CoG, many of them will point to fossil evidence as supposed evidence that it did occur, regardless of defining the mechanisms as to how it might have occurred.
Supposed Fossil Record Evidence
Many evolutionists will point to so-called dinosaurs in the fossil record as supposed evidence of dinosaur-to-bird evolution, including evidence for the purported anterior shift in CoG and the evolution of other flight characteristics. Evangelista (2013) conducted in-depth aeronautical research regarding the flight dynamics of birds that includes numerous calculations for time-dependent CoG/CoM, pitch stability, roll stability, and yaw stability. Evangelista (2013, 61) includes a section for “tests in dinosaurs,” but notes the following:
I report the effects of posture and morphology on the static aerodynamic stability and control effectiveness of physical models based loosely on a feathered dinosaur, Microraptor gui, from the Cretaceous of China.
However, Leinweber (2024) makes a strong case that Microraptor gui should be reconstructed to look much like a bird rather than a dinosaur with feathers. Extinct birds have an anterior CoG, as expected, to enable flight. Although the debate about Microraptor gui’s classification and reconstruction is likely to continue, its possible reconstruction as a bird should be taken into consideration when considering CoG claims. Evolutionary researcher Burnham (2008, 38) acknowledges Microraptor’s apparent anterior CoG/CoM:
It is apparent from the skeletal construction of Microraptor that this skeletal anatomy was powered by a strong muscular system in the chest and upper arms. In fact, the body outline in the pectoral area and forelimbs far exceeds that of the pelvic area and hindlimbs . . . The glenoid was positioned high on the back and forward. This places the CoG in a position giving the animal a high center of mass at the shoulders.
Extinct birds with functional flight design having an anterior CoG close to their center of lift is no surprise from a creation perspective, though there are two possible objections to consider.
First, if any flying creatures are found with a CoG that does not align anteriorly with the wings, then it’s possible that the creature has a second set of lift-generating wings in a different location on its body. In this case, the calculated neutral point position would need to account for pitching moments from both sets of wings, and that position would still align closely with the CoG. In other words, in a multi-wing system, the neutral point may fall somewhere between the two sets of wings, but for pitch stability, the CoG still must be at or forward of the neutral point. Some creatures such as Microraptor had feathers on their legs, which some evolutionary researchers claim to be a second set of wings. Dyke et al. (2013) call Microraptor a “feathered dinosaur” and this belief leads to hypotheses and assumptions such as primitive flight ability and lack of a specialized and dedicated flight control system. Dyke and colleagues and other researchers such as Chatterjee and Templin (2007) conducted aerodynamic calculations and conclude that Microraptor’s flight abilities were limited to gliding, but this too may be inaccurate and based on evolutionary assumptions. Evolutionary assumptions generally do not account for this creature’s birdlike morphology, such as shoulder mobility and flight muscles consistent with wing flapping ability. Dyke and colleagues calculate a CoG range that is somewhat anterior, but with the most posterior end of the range being close to the base of the tail. Evolutionary researchers may point to this research and attempt to claim that hindlimb feathers might have compensated for a more posterior CoG, thus enabling gradual changes to support other body modifications, but Leineweber (2024, 44) notes the following problem regarding the hindlimb models:
Despite these claims, others have concluded that Microraptor’s sprawled hindlimb model was based on poor data (Brougham and Brusatte 2010), and its range of motion would not support sprawling or flapping hindlimbs (Manafzadeh and Padian 2018). Certainly, God may have designed Microraptor’s hindlimb feathers to contribute to its agility in flight, but comparing with similar existing birds will likely be the most informative approach, as argued above. A sprawling hindlimb would diverge from extant birds which, as discussed, use soft tissue anatomy to keep their knees tucked in a crouched position and so maintain balance while walking.
Moreover, a likely more accurate reconstruction of Microraptor such as Leineweber’s may enable future creationist research to calculate a more anteriorly-placed CoG range compared to that of Dyke and colleagues, as well as aerodynamic calculations consistent with powered flight. Other evolutionary researchers conclude an anterior CoG for Microraptor as noted above. Therefore, pointing to Microraptor as a supposed transitional form for CoG shift is not well supported.
The second caveat to extinct birds having an anterior CoG is the debated location of the CoG for Caudipteryx. Jones et al. (2000, 716) state the following:
Surprisingly, Caudipteryx, described as a theropod dinosaur, possessed an anterior centre of mass and hindlimb proportions resembling those of cursorial birds.
However, some researchers such as Talori et al. (2019, 2) disagree, and note that its CoG “differs” from modern birds:
Caudipteryx further differs from modern birds which have abbreviated tails and forward centered mass locating near the wings. However, the most primitive winged dinosaur, Caudipteryx, is clearly terrestrial, investigating the aerodynamic properties of the proto-wings of Caudipteryx has the potential to shed light on the origin of avian flight.
The label “primitive winged dinosaur” is based on evolutionary presuppositions. Perhaps Caudipteryx was indeed terrestrial, but as a flightless bird, and not a dinosaur as noted by Geist and Feduccia (2000). Therefore, rather than shedding light on supposed dinosaur-to-bird evolution, studying Caudipteryx may simply enable researchers to gain a deeper understanding of God’s design for this creature. More research might be needed to pin down the most likely location for its CoG, but if this creature is indeed a flightless bird, then there are two possibilities from a creation perspective: God designed it with a morphology suited for walking and running instead of flying, right from the beginning, or this creature lost flight ability from an originally flight-capable kind via loss of genetic information.
As noted in the Methodology section, this portion of the paper is not meant to be an exhaustive analysis of evolutionary claims regarding the fossil record. Some researchers such as Allen et al. (2021) used statistical analysis and cladistics to support their claim that dinosaur-to-bird evolution involved a gradual transition from hip-based to knee-based locomotion. Creationist researchers such as Junker (2024) have strongly critiqued similar cladistical analyses for numerous other traits such as feather types and respiratory systems. Future creationist research can critically examine evolutionary cladistical analyses pertaining to flight and locomotion.
Predictions
The Creator God of dinosaurs, birds, and everything else in creation states the following in His Word:
For not all flesh is the same, but there is one kind for humans, another for animals, another for birds, and another for fish.—1 Corinthians 15:39 (ESV)
This Scripture implies a significant difference in physical nature (“flesh”) between land animals (“another for animals”) and birds (“another for birds”). Flying creatures were created on Day Five of Creation Week (Genesis 1:20–21), and land animals, such as dinosaurs, were created on Day Six of Creation Week (Genesis 1:24–25). Since flying creatures were designed for flight, and dinosaurs were designed for walking/running, this author makes several predictions regarding future research and fossil record findings.
First, ongoing and future creationist research, free from evolutionary assumptions, will continue to make a distinction between extinct birds and dinosaurs found in the fossil record. This research will continue to refute the claim that dinosaurs evolved into birds, and that some dinosaurs were able to fly. One caveat to this prediction is that some professed creationist researchers may obtain faulty conclusions in their research if it is not completely free from evolutionary assumptions.
Second, evolutionary researchers might continue to label extinct birds as dinosaurs as supposed evidence for an evolutionary, anterior shift in CoG. Nevertheless, such claims might be refuted with the support of ongoing and future creationist research that refutes evolutionary cladistical claims and analyses, while continuing to draw a line of distinction between birds and dinosaurs in the fossil record.
Third, additional fossil finds of extinct birds that were designed for flight will always appear to have a body shape conducive to an anterior CoG that is closely aligned to the center of lift of the wings, with one caveat and one possible exception. The caveat is if God designed a bird kind to be flightless from the beginning, then it may have a posterior CoG, so this prediction is limited to extinct birds designed for flight (that is, extinct flight-capable birds will consistently reflect flight-ready design). The exception to this prediction is if any flying creatures are found with a CoG that does not align with pectoral wings, then it’s possible that the creature has a second set of lift-generating wings in a different location on its body. In this case, the calculated neutral point position would need to account for pitching moments from both sets of wings, which would still align closely with the CoG. Such a finding would still fit with the creation model and would not require an evolutionary interpretation.
Fourth, evolutionary researchers might continue to attempt explanations to demonstrate the feasibility of evolutionary, anterior shift in CoG, but such explanations are expected to fall short of observational science (not directly observable or experimentally demonstrable) due to two key constraints. First, such explanations may ignore genetic constraints, such as the inability of genomes to add new, useful genetic information necessary to add new, complex functionality. Second, such explanations may gloss over the need for irreducibly complex functions to evolve simultaneously.
Future Research
This author recommends several topics for future research.
Sanders (2025) challenged many of the evolutionary claims regarding dinosaurs with feathers. Additional research may further define feathers as being a distinct characteristic to birds. Since many evolutionary researchers believe that dinosaur tails shortened, this topic of research is necessary to determine if dinosaur tails ever had feathers. If dinosaur tails are increasingly found to be featherless, then this confirms an insurmountable obstacle for dinosaur-to-bird tail evolution: adding new genetic information for a featherless tail to grow feathers, let alone the necessity for dinosaurs to also evolve feathers on their forelimbs.
A more accurate reconstruction of Microraptor such as Leineweber’s may enable future creationist research to calculate a more anteriorly-placed CoG range compared to that of Dyke et al. (2013). Creationist researchers can redo the aeronautical experiments and calculations of that of Dyke and colleagues by constructing a model based on Leineweber’s reconstruction (2024), which may yield aerodynamic calculation results that are consistent with powered flight, rather than limited flight such as gliding.
Creationists can also conduct experimental studies on models that contrast pitching moment numerical values between birds and hypothesized transitional forms, physically demonstrating the pitch instability differences.
Allen and colleagues (2021) used statistical analysis and cladistics to make the case that dinosaur-to-bird evolution involved a gradual transition from hip-based to knee-based locomotion. Creationist researchers can critically examine these claims with similar rigor to that of Junker’s (2024) analysis. Evolutionary cladistics are wrought with assumptions that blur the lines between dinosaurs and birds, and as Junker demonstrated, oftentimes the cladistics don’t quite line up with the fossil findings. Allen and colleagues’ claim of gradual transition from hip-based to knee-based locomotion might exhibit the same problems under similar scrutiny.
Fossilized remains of creatures such as Caudipteryx continue to be hotly debated. Further research may confirm their status as flightless birds, and such research may help creationists understand if God created them to be flightless or if they lost flight ability through mutations.
Conclusion
God created the flying creatures on Day Five of Creation Week (Genesis 1:20–21), and land creatures on Day Six (Genesis 1:24–25). When aeronautical engineers design airplanes so that the CoG is near (usually slightly forward of) the neutral point and center of lift, they are utilizing principles that God used in His design for flying creatures, in accordance with the laws of physics that He established. Flight stability in both aircraft and animals requires similar CoG placement. While evolutionists often conflate birds with theropod dinosaurs or theropod dinosaurs with birds in the fossil record due to their evolutionary presuppositions, creationists can expect to find distinct differences between the two types of creatures when analyzing both extant birds and fossil evidence. When stripping away evolutionary assumptions and interpretations, current CoG findings seem to be consistent with these expected differences: an anterior CoG for flying creatures (close to center of lift), and a posterior CoG for theropod dinosaurs (close to the hip joint). God designed flight-capable birds for flying, and dinosaurs for walking. Future creationist research is expected to confirm these observations, and this paper recommends further research because the aeronautical perspective may help settle the debate when combined with the perspectives of paleontology, biomechanics, and biology. Since many evolutionists effectively claim that dinosaurs achieved a similar feat as that of the Wright Brothers, aeronautical considerations such as CoG are worth exploring within the dinosaur-to-bird evolution debate.
References
Allen, Vivian, Karl T. Bates, Zhiheng Li, and John R. Hutchinson. 2013. “Linking the Evolution of Body Shape and Locomotor Biomechanics in Bird-Line Archosaurs.” Nature 497, no. 7447 (2 May): 104–107.
Allen, Vivian R., Brandon M. Kilbourne, and John R. Hutchinson. 2021. “The Evolution of Pelvic Limb Muscle Moment Arms in Bird-line Archosaurs.” Science Advances 7, no. 12 (19 March): eabe2778.
Bramwell, Cherrie D., and G. R. Whitfield. 1974. “Biomechanics of Pteranodon.” Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 267, no. 890 (July 11): 503–581.
Burnham, David A. 2008. “A Review of the Early Cretaceous Jehol Group of Northeastern China and a Revision Concerning the Origin of Flight Paradigm.” Oryctos 7 (January): 27–43.
Carrano, Matthew T, and Andrew A Biewener. 1999. “Experimental Alteration of Limb Posture in the Chicken (Gallus gallus) and Its Bearing on the Use of Birds as Analogs for Dinosaur Locomotion.” Journal of Morphology 240, no. 3 (June): 237–249.
Carrano, Matthew T. 1998. “Locomotion in Non-Avian Dinosaurs: Integrating Data from Hindlimb Kinematics, in Vivo Strains, and Bone Morphology.” Paleobiology 24, no. 4: 464. http://www.jstor.org/stable/2401181.
Chatterjee, Sankar, and R. Jack Templin. 2007. “Biplane Wing Planform and Flight Performance of the Feathered Dinosaur Microraptor Gui.” Proceedings of the National Academy of Sciences of the United States of America 104, no. 5 (January 30): 1576–1580.
Cserhati, Matthew. 2023. “Feathered Turtles?” Answers Research Journal 16 (September 27): 491–499. https://answersresearchjournal.org/dinosaurs/feathered-turtles/.
Degernes, Laurel A., and Alan Feduccia. 2001. “Tenectomy of the Supracoracoideus Muscle to Deflight Pigeons (Columba Livia) and Cockatiels (Nymphicus Hollandicus).” Journal of Avian Medicine and Surgery 15, no. 1 (March): 10–16.
Dyke, Gareth, Roeland de Kat, Colin Palmer, Jacques van der Kindere, Darren Naish, and Bharathram Ganapathisubramani. 2013. “Aerodynamic Performance of the Feathered Dinosaur Microraptor and the Evolution of Feathered Flight.” Nature Communications 4 (18 September): Artice no. 2489.
Evangelista, Dennis José. 2013. “Aerial Righting, Directed Aerial Descent, and Maneuvering in the Evolution of Flight in Birds.” Ph.D. diss, University of California. https://www.proquest.com/openview/1170fe5bc190cf0d68887ceec7328a1f/1?pq-origsite=gscholar&cbl=18750&diss=y.
Farlow, James O., Stephen M. Gatesy, Thomas R. Holtz Jr., John R. Hutchinson, and John M. Robinson. 2000. “Theropod Locomotion.” American Zoologist 40, no. 4 (August): 640–663.
Gatesy, Stephen M. 1990. “Caudofemoral Musculature and the Evolution of Theropod Locomotion.” Paleobiology 16, no. 2: 170–186.
Geist, Nicholas R., and Alan Feduccia. 2000. “Gravity-Defying Behaviors: Identifying Models for Protoaves.” American Zoologist 40, no. 4 (August): 664–75.
Grossi, Bruno, José Iriarte-Díaz, Omar Larach, Mauricio Canals, and Rodrigo A. Vásquez. 2014. “Walking Like Dinosaurs: Chickens with Artificial Tails Provide Clues about Non-Avian Theropod Locomotion.” PLoS ONE 9, no. 2 (February 5): e88458.
Hartman, Scott. 2020. “Theropod Skeletal Reconstructions.” Dr. Scott Hartman’s SkeletalDrawing.com. https://www.skeletaldrawing.com/theropods/sinosauropteryx.
Haynes, Gabriela. 2022. “The Debate over Classification of Archaeopteryx as a Bird.” Answers Research Journal 15 (September 14): 285–300. https://answersresearchjournal.org/dinosaurs/debate-classification-archaeopteryx-bird/.
Haynes, Gabriela. 2023. “Reply to the “Response to ‘The Debate over Classification of Archaeopteryx as a Bird’”.” Answers Research Journal 16 (December 6): 595–634. https://answersresearchjournal.org/dinosaurs/reply-response-debate-classification-archaeopteryx-bird/.
Hutchinson, John R., and Vivian Allen. 2009. “The Evolutionary Continuum of Limb Function From Early Theropods to Birds.” Naturwissenschaften 96, no. 4 (24 December): 423–448.
Jones, Terry D., James O. Farlow, John A. Ruben, Donald M. Henderson, and Willem J. Hillenius. 2000. “Cursoriality in Bipedal Archosaurs.” Nature 406, no. 6797 (17 August): 716–718.
Junker, Reinhard. 2024. “Are Birds Dinosaurs? A Critical Analysis of Fossil Findings.” Answers Research Journal 17 (QApril 24): 223–309. https://answersresearchjournal.org/dinosaurs/are-birds-dinosaurs/.
Leineweber, Joel. 2024. “Microraptor Reconstructed as a Bird.” Answers Research Journal 17 (May 15): 317–352. https://answersresearchjournal.org/dinosaurs/microraptor-reconstruction/.
Lisle, Jason. 2018. “Refuting Dubious Claims Regarding Natural Selection.” Answers Research Journal 11 (December 5): 301–340. https://answersresearchjournal.org/refuting-dubious-natural-selection/.
Nelson, Robert C. 1998. Flight Stability and Automatic Control. 2nd edn. Boston, Massachusetts: WCB McGraw Hill.
Paul, Gregory S. 2016. The Princeton Field Guide to Dinosaurs. 2nd ed. Princeton, New Jersey: Princeton University Press.
Rashid, Dana J., Susan C. Chapman, Hans C. E. Larsson, Chris L. Organ, Anne-Gaelle Bebin, Christa S. Merzdorf, Roger Bradley, and John R. Horner. 2014. “From Dinosaurs to Birds: A Tail of Evolution.” EvoDevo 5 (29 July): Article no. 25.
Sanders, Harry III. 2025. “Biochemistry of Dinofuzz: Feathers, Filaments, Fuzz, or Folly?” Answers Research Journal 18 (March 5): 125–131. https://answersresearchjournal.org/biochemistry/biochemistry-dinofuzz-feathers-filaments-fuzz-folly/.
Talori, Yaser Saffar, Jing-Shan Zhao, Yun-Fei Liu, Wen-Xiu Lu, Zhi-Heng Li, and Jingmai Kathleen O’Connor. 2019. “Identification of Avian Flapping Motion from Non-Volant Winged Dinosaurs Based on Modal Effective Mass Analysis.” PLOS Computational Biology 15, no. 5 (May 2): e1006846.
Tecuceanu, Cristian, Dumitru Popovici, Mariana Popovici, and Peter Kalmutchi. 2023. “Mechanical Strength of the Aircraft Control Chains in Certain Unusual Cases.” Incas Bulletin 15, no. 4 (December): 247–262.
Tan, Change Laura. 2022. “The Absence of Universally Conserved Protein-Coding Genes.” Answers Research Journal: (April 20) 83–95. https://answersresearchjournal.org/genetics/absence-protein-coding-genes/.