The views expressed in this paper are those of the writer(s) and are not necessarily those of the ARJ Editor or Answers in Genesis.
Abstract
God desires to reveal His character enough that He does not keep Himself from being known by His creation. He allows us to know Him, as He reveals truths about His character and image. Knowing God is the most important duty we can have. God makes Himself known to His creation through His Scriptures and His world. As biologists, we get to see His world in an amazing and enlightening way. As we are confronted by God’s creation, we can be confused by aspects of His creation that do not function well. Parasites don’t seem to fit into a design that God would call “good.” Organisms, that function as parasites now, may have been created to function as part of the body of the host, for mechanisms of gene transfer, and/or as modes of selective pressure on the host. This study focuses on raccoon roundworm (Baylisascaris procyonis), which seems to be an important part of individual raccoon behavior and raccoon population genetics, and lend support to these hypotheses. As we continue to learn more about parasites, we will understand better how the parasitic lifestyle resulted from the Fall.
Keywords: Genesis, biology, creation, symbiosis, parasitism, parasites
Introduction
Attaining knowledge
It is important to remember that God has created His world to be known (Psalm 19). Epistemology is concerned with two aspects of knowledge: the nature of knowledge and how it is attained. Each discipline aims to establish its own epistemological framework, describing the nature and acquisition of knowledge differently. As Christians, we must remember that the Scriptures are the final authority in faith and life, and are sufficient in all areas addressed by the biblical authors (Anonymous 2013; Proverbs 1:7). The Scriptures are the foundation upon which all other disciplines are built and assessed. They are the very Word of God, spoken to man, and profitable in all areas of life (2 Timothy 3:16–17; 2 Peter 1:19–21). What does Scripture have to say about knowledge?
Scripture describes knowledge as being inseparable from God (Job 28). The first instance of a word translated as knowledge is in Genesis 2. Genesis 2 describes the immanence of God as He created. It describes a tree of the knowledge of good and evil. God forbids Adam from eating the fruit of this tree (Genesis 2:17). This passage, and the passage of the Fall in Genesis 3 indicate that knowledge is associated with understanding. Sometimes knowledge can be a dangerous influence that needs to be handled well. Adam desired this knowledge outside of God’s time and design, bringing death into the world (Romans 5:12–19). During Zechariah’s prophecy in Luke 1:67–79, the Holy Spirit expanded on the truth that knowledge is a type of understanding. He taught that John the Baptist would prepare the way for the Lord by giving the “knowledge of salvation to his people” (Luke 1:76–77 ESV). As the verse unfolds, we see that this knowledge is a prerequisite for the forgiveness of sin. In this sense, knowledge, coupled with God’s grace, has the power to remove the bondage of sin (Romans 6:18). The Scriptures speak much more about the nature of knowledge, but it usually seems to be associated with a knowledge of and relationship with God (Job 28:28; Psalm 111:10; Proverbs 1:7; 9:10; 15:33), His creation and our role as children in His creation.
The main focus and purpose of life is to know God personally (Packer 1973). Jesus described eternal life as the knowledge of God and Jesus Christ (John 17:3). In this life, the thing that brings the most happiness and contentment is the knowledge of God (Jeremiah 9:23–24). Knowledge of God is the unifying principle of all knowledge (von Rad 1972, p. 68). As such, God finds pleasure more in our knowledge of Him than anything else (Hosea 6:6). Attaining knowledge of the creation should ultimately point us back to God.
The first step in the acquisition of knowledge is to learn where to seek. The Apostle Paul encourages us to filter our thoughts and confront errant ways of thinking to better serve our Lord Jesus Christ (2 Corinthians 10:5). The first step in this development is to recognize that all knowledge is a single entity that comes from the Creator (Dockery 2008). Accordingly, all truth comes from God, who is a “single universe of knowledge” (Dockery 2008, p. 46). We serve a God who is truth and delivers that truth to His creation. How does our God reveal knowledge?
The second step is to know how to seek knowledge. There are “secret things” that belong to the LORD, but He reveals other things that we may obey Him (Deuteronomy 29:29). Seeking knowledge from God requires listening to His Word (Packer 1973; Deuteronomy 4:10; 17:19; 31:12–13). Knowledge is attained as we discover these things that God reveals to us. Packer (1973) demonstrates that God reveals aspects of His nature and character to us, and we must accept His invitations to attain this knowledge. God’s special revelation comes from His Word. His general revelation is expressed in His creation.
God has revealed and continues to reveal Himself as He chooses to sinful men through His Holy Scriptures (Morris 1976). God chooses when and how to reveal truth through His Word. As Morris (1976) points out, special revelation has taken place through the history of the Hebrews. The Old Testament records literal history, but only the history of God’s redemptive plan through the Hebrews. Special revelation reaches its climax in the life, death, and resurrection of Jesus Christ, but continues through the culmination of God’s redemptive plan. Each and every word of Scripture reveals some of God’s truth. This truth seems to focus on the story of God redeeming man from the effects and consequences of Adam’s sin (Romans 5). Scripture is the source of revelation that provides the rubric used to assess all other knowledge. In other words, special revelation makes other forms of revelation possible to interpret.
The Creator uses His creation as evidence of His existence (Morris 1976). Morris (1976) argues that a reflection on creation reveals not only that God exists, but also that He is a particular type of God. Psalm 19:1 (ESV) declares, “The heavens declare the glory of God, and the sky above proclaims his handiwork.” Psalm 19 continues to describe this revelation. Creation declares the existence of God, by displaying His handiwork. While there are no words spoken, creation still “pours out speech,” and “reveals knowledge” all day long (Psalm 19:2 ESV). Paul declares that God’s power and deity can be seen in His creation (Romans 1:20). Paul and Barnabas state that God’s general grace to all bear witness about His existence (Acts 14:17). The fact that creation still functions and that man can and does subdue it is a reflection of the existence of God. God created man in His image (Genesis 1:26). Even fallen man still demonstrates the image of God (Genesis 5:3). Though fallen, this image is powerful. It allows man to exercise power over creation as God’s vice-regent over creation. This function is facilitated by the fact that man is made in God’s image and likeness. When man exercises this ability, he knowingly, or unknowingly, demonstrates the existence of a God more powerful than creation.
It is important to remember that the very nature of revelation is something that is revealed by God to certain individuals (Morris 1976). Therefore, God’s general revelation is not open to differing interpretation by all individuals. It must be brought from God to an individual. God must disclose it in His grace, and does so for a purpose. Does this mean that believers see a different physical universe than nonbelievers? This is certainly not the case. Christians see what everyone perceives, but also see what God is declaring through His creation (Boyd 2008, p. 171; Morris 1976).
Purpose and evaluation of acquired knowledge
The purpose of attaining knowledge is to become wise as God is wise. True wisdom is “the power to see, and the inclination to choose, the best and highest goal, together with the surest means of attaining it” (Packer 1973, p. 90). Wisdom is so much more than knowledge, but it requires the knowledge of God, His creation and our role to be correct. Longman’s comments on Job 28:28 reinforce this point further when he describes the verse describing the need to fear the Lord to gain the wisdom that only God possesses otherwise (Longman 2012, p. 333).
All knowledge must be evaluated by God. Therefore, all knowledge acquired through reading God’s Word and studying God’s creation must be checked by God Himself. The Holy Spirit interprets Scripture and allows us to receive it (Packer 1973). We cannot recognize the truth of Scripture apart from the work of the Spirit (Morris 1976). Jesus tells His disciples that the Spirit will teach them and remind them of His teachings (John 14). He also teaches His disciples that the Spirit will convict the world of sin, righteousness, and judgment (John 16). The Spirit makes Scripture and creation able to be understood, and convicts men of these truths. Morris (1976) argues that apart from the working of the Spirit, the truths of Scripture become something that the believer must defend. He argues that true revelation has the power to change men into believers, not militant defenders of truth. We can only attain knowledge that God reveals and interprets. Apart from this, knowledge is incomplete at best, and significantly distorted (Romans 1:21–25; 1 Corinthians 1:18–2:5).
God Creates
Empiricism vs integration
The dominant worldview of scientists is empiricism. The empirical worldview holds that all concepts and knowledge available to humans must rely on empirical evidence. The observer must be able to experience the knowledge with a sensory experience. According to this worldview, any idea or belief that is not grounded in empirical evidence is empty and meaningless. Stephen Jay Gould argued that the only way to have religion and science coexist is to let science have the facts and religion have the values (Bloom 2005). Bloom (2005) attempts to reduce all knowledge to scientific knowledge, by explaining the existence of religion in scientific terms. From his perspective, and the perspective of all empiricists, religion has no place in scientific discussions. In order to be a scholar in the sciences, one would have to be a practical atheist, believing that there is a god, and that he is not necessary for the explanation of life.
This prevailing worldview is not consistent with integrating Scripture and faith into the discipline of biology. From this perspective, Scripture can have no say on why or how we study biology. This is the biggest obstacle to the integration of faith and learning in biology. Naturalistic thinking is a part of almost every aspect of biology; accordingly, the integration of faith and learning in biology will require a completely different approach to studying life.
Scripture is clear that Christians must have a mind that is qualitatively different than the minds of the unredeemed (Romans 12:2; Ephesians 4:17). This requires Christians to study life differently. A truly Christian perspective on biology must hold to certain presuppositions about life: God created and is actively involved in the natural world, and reveals Himself through His creation; humans are image bearers of God that are affected by sin, but must still exercise stewardship over God’s creation; and the Scriptures provide information about the physical universe and undergird all discoveries in biology (Jones 2013). What does Scripture have to say about biology?
Biology and special revelation
Scripture speaks to two aspects of biology more than any other. The first aspect speaks about how we study biology. The second speaks to why we study biology. We must study biology from the perspective that God is the source of life (Genesis 1–2). We study biology because we are required to steward creation (Genesis 1:26–28).
Scripture begins at creation. “In the beginning, God created the heavens and the earth” (Genesis 1:1 ESV). A literal reading of Genesis 1 presents that God created the day and night on Day One of creation, followed by five more days of creation that culminate in His creation of man after His own image. Scripture declares that all life has its origin in God. Genesis expands on the creation of God to demonstrate His immanence and intimacy with creation. Colossians 1:17 goes on to demonstrate that God is still involved in creation. Creation is not a single historical event, but continues through the handiwork and sustenance of God (Butkus 2002). God’s handiwork is so intrinsic to creation, that His power and attributes can be seen throughout creation (Psalm 19; Romans 1:20). We study biology in light of what Scripture teaches about origins. Therefore, the questions we ask, and hypotheses we make need to be in this context. If God continues to sustain and uphold His creation, there will not always be a physical explanation for life.
Scriptural truths on why we study biology are an important aspect of the integration of faith and biology. Man is to steward creation. The very first command given to man in Scripture is to multiply, fill the earth, subdue it, and exercise dominion over it (Genesis 1:28). Man was created to represent God to creation. The word for dominion, rādāh, implies a kingship role (Butkus 2002). Since it is an imperative, it conveys the sense of “Rule!” Man is to exercise dominion over creation the way a king would exercise dominion over his kingdom. If man is to live up to His Godly image, this dominion should be aimed towards the flourishing of creation. The next image of stewardship displays man working in the garden and keeping it (Genesis 2:15). There are 24 more explicit references in Scripture for stewardship, and many more passages that reveal the need to steward creation (Butkus 2002). Psalm 8 serves to link God’s handiwork displayed in His creation with man’s obligation to keep that which God has put him in charge. It is a humbling task to care for the creation that displays the handiwork of God. In order to do it well, we must expand our understanding of the creation.
Biology and general revelation
God reveals many aspects of His nature through His creation. The non-physical, non-material aspect of life illustrates God’s spiritual life (Wise 2013). Wise (2013) explains how several more biological phenomena illustrate aspects of God’s nature. The beauty in creation illustrates God’s glory and His care for His creation. The required complex cycles of essential nutrients represents God’s love and provision for His creation. The biological systems that can be found in every living organism illustrate God’s unity. All life has personality, illustrating God’s personhood. Mutualism illustrates God’s goodness and relation to His creation. Complexity illustrates God’s omniscience. The existence of language illustrates God is an intelligent being that desires to communicate with His creation. God has revealed and continues to reveal Himself through His creation. When we study creation, we develop knowledge of the Creator that helps us serve Him better.
It is important to remember that God has created His world to be known (Psalm 19). The Bible speaks in complementary terms about the biological inquiries of Solomon, as he spoke about all sorts of plant and animal life (1 Kings 4:33). As we learn more about creation, we become better stewards of it, and develop a fuller appreciation for how God has created. As one studies creation, it becomes obvious that God did not need to specially create every single species (Wood and Garner 2009). He created animals in “Genesis kinds,” with genetic diversity, capable of limited speciation (Wood and Garner 2009, p. 3). Scripture does not require species to be fixed, but rather implies that created kinds are dynamic (Garner 2009). How then do Genesis kinds diversify?
Symbionts live alongside, and often inside other animals (hosts). These symbionts allow the host to interact more intimately with its environment (Francis 2013). Symbionts, especially parasites, exert enormous pressure on host populations to change (Ingle 2013). This pressure, coupled with other mechanisms can generate new species (Francis 2009). God created animals to relate in very intimate ways with the environment. This relationship may have fueled much of the diversification within a Genesis kind. God created animals to fill the earth (Genesis 1:22). In order for animals to do this, they must diversify as they enter new environments. As we understand creation better and how animals diversify and relate to other creatures, we can become better stewards of creation. We can help creation flourish the way God intended. How then should we address and think about symbiotic organisms that reduce the fitness of their host, namely the parasites?
Parasitism in God’s Creation
Complications with parasites being “good”
Genesis 1 describes the creation of the heavens and the earth by God in a single week. In Genesis 1:26 we see the first instance of relationship when God describes His own relationship within the Trinity, “Let us make mankind in our image, in our likeness” (NIV, emphasis added). This is the first time relationship is introduced in Scripture. God relates with Himself in the different persons of the Trinity, but He also relates with His creation. We are exposed to His immanence in verses 26–30 as He creates man in His own image, with His own character, and then even relates to man, giving him purpose. Scripture reveals here that man is meant to relate to the rest of creation by subduing it and ruling over it. Within the context of these relationships, Genesis 1 also reveals an intimate relationship between plants and man (verse 29) and plants and animals (verse 30), as plants provide food for the rest of creation. Relationship seems to be a fundamental biblical concept from the very first chapter of Scripture (Francis 2009). On the surface, the relationship between plants and animals is completely one-sided. Animals benefit at the expense of the plants. Could there have been animals relating with animals in similar one-sidedness in God’s original creation? In other words, does parasitism fit within God’s original design and intent of His creation?
Parasites often invoke thoughts of disgust and sometimes panic. These aren’t the thoughts that are typically associated with the reading of God’s creation as declared “good” throughout Genesis 1. Instead, parasites are creatures that benefit at the expense of their host, often living amongst the host tissues and evading host immunity. It is difficult to reconcile this kind of existence with a benevolent Creator, who would declare this to be good. Charles Darwin struggled with the possibility of a loving and omnipotent God creating Ichneumonid wasps (parasitoid wasps) that deposit their larvae inside or near a living animal that is then eaten alive by the larvae (Dawkins 1976, p. 67). It seems that parasites must have been designed to do something besides reduce the fitness of their respective hosts, so what might this design have been?
Changing perspectives
McElroy and de Buron (2014) found that in 58 out of 101 studies (58%) parasites reduced host performance capacity (how well an organism can execute a given behavior or ecologically relevant task). While host age and location of infection impacted the reduction in host performance capacity, it is important to note that many parasites cause significant reduction in host fitness. However, the pathological effects of parasites are often dependent on a number of factors, with many parasites behaving more like commensals (organisms that benefit from the host but not at the expense of host fitness). For instance, Granath (2014) found a significant reduction in the intensity of Myxobolus cerebralis in Salmonid fish and the corresponding pathology after the restoration of Upper Willow Creek in west-central Montana. Granath argued that habitat rehabilitation reduces abundance of parasites by reducing transmission of the parasites among hosts. Even in symbiotic species that do lower host fitness, the pathology associated with the presence of the parasite is often minor. Furthermore, parasite-induced mortality is rare in hosts. Ungulates are common hosts for the lancet liver fluke (Dicrocoelium dendriticum), with fluke-induced mortality often restricted to older hosts with already reduced fitness (Beck et al. 2014). One factor that can increase the pathological effects of a parasite is infestation rate. Potential hosts often adopt behaviors that minimize exposure to parasite individuals and thus reduce infestation rates. Goitered gazelles (Gazella subgutturosa) reduce group sizes during the breeding season of the parasitic botfly (family Hypodermatidae) to keep infestation rates low and thus reduce pathologic potential (Blank and Yang 2014).
Parasitism represents the most common consumer strategy on earth (Poulin and Morand 2000), and to ignore the conservation of parasitic organisms is to ignore the conservation status of the majority of life on earth (Gómez and Nichols 2013). Parasites are an often overlooked and underrepresented component of food webs, but often have incredible impacts on the structure and function of these systems (Rossiter 2013). Rossiter (2013) shows how parasites can be added to models of food webs in a simple, accurate, and helpful way. Gómez and Nichols (2013) found several dozen species of parasites that are critically endangered or extinct. It is currently thought that we are losing many species of parasites with the extinction of free-living hosts of the parasites (Dunn et al. 2009). Gómez and Nichols (2013) contend that we should take the conservation of these organisms seriously. Given the variety and richness of parasite species, they must have been part of God’s design. Therefore, I can’t help but agree with Gómez and Nichols, but for a different reason. I believe that the proper stewardship of creation requires that we help creation to function better than it currently does and to support biodiversity when possible. A more complex and robust creation better represents the complexity and beauty of the Creator (Psalm 19). To do this, we must answer the following question: What might the design of parasites have been, and how can it be restored?
Three ways of thinking about parasites
It is important to note that views on what animals that currently live as parasites may have been designed to do are not exhaustive nor are they mutually exclusive. I believe that all three are part of the origin design of symbiotic organisms in general, and parasitic organisms specifically. The original animals living inside of other animals as part of God’s design for His creation could have been part of the holobaramin of the host, they could have been designed as a mechanism of horizontal gene transfer, and they could have also been designed to provide a mechanism for selection as animals diversified and adapted to different ecosystems. A holobaramin is a complete set of organisms that belongs to a single baramin (created kind).
Francis (2009) asked whether we should consider microbial symbionts whether they live inside or outside the host as part of the holobaramin of the host. He argued that these symbionts could be thought of as a body part or tissue in the original baramin, as long as they do not contribute to extensive morphological change when present in the host. Animal parasites can also be thought of in this context. While significantly complex at the tissue and organ level, some animals live now as obligate parasites using a single animal species as the definitive host. Raccoon roundworm, Baylisascaris procyonis, is an intestinal nematode that parasitizes raccoons, Procyon lotor. Raccoon roundworm prevalence (the proportion of individual hosts in a sample that are infected with the parasite) often exceeds 70% (Ingle et al. 2014). Raccoon roundworm cannot fully develop outside of raccoons. This obligate parasite causes no pathology in raccoons unless there are several dozen present. Given how prevalent this animal is inside of raccoons and the lack of pathology when it is present in low numbers, could we consider it to be a multicellular, genetically distinct part of the holobaramin of the host? Animals such as these could have been designed to be a portion of the host’s body that interacted more intimately with the environment by spending a portion of its life history outside of the host. Do only the cells that originate from the zygote count as part of the baramin? Raccoon roundworm prevalence does seem to correlate with individual raccoon behavior and raccoon population dynamics (Ingle 2013).
Most creation biologists recognize the existence of speciation, and the need for rapid speciation and diversification following the global Flood introduced in Genesis 6. The magnitude of this diversification would have been so extreme that we infer that nothing like it is taking place now and diversification has ceased (Wood 2002). What could have provided a mechanism for such rapid and intense diversification? Wood (2002) advocates for mobile genetic elements, that he calls Altruistic Genetic Elements (AGEs), being the mechanism behind this rapid speciation. These transposable and mobile sections of DNA could provide the rapid and permanent change (as long as the element does not subsequently leave the genome of the animal) necessary for the diversification and reproduction isolation of the baramins coming off the Ark. This proposed mechanism for genetic and phenotypic diversification requires some way for the AGEs to make it into the genome of the target baramin as it moves into a new environment, spreading out from the Ark. Wood (2002) argues that these AGEs could explain convergence and coevolution when animals are in similar ecosystems, or a symbiotic relationship respectively. Zhang et al. (2012) found evidence that microRNAs from plants are present in human serum and down-regulate the expression of low-density lipoprotein receptor adapter protein 1 (LDLRAP1) expression in liver tissue. The decrease in LELRAP1 expression decreases LDL removal from mouse plasma. These microRNAs are acquired orally through food intake. Comparatively, many parasites, especially in Clade Neodermata have complex life cycles that require the parasite to move through multiple hosts to complete their life cycle. Could this be a relic of God’s creation to introduce AGEs to animals living in the same ecosystem? Raccoon roundworm, as mentioned above, also typically requires at least one intermediate host before finishing development in raccoons. This parasite is virtually ubiquitous in raccoons (with the exception of the southeastern USA), and makes a good candidate for having been designed to deliver AGEs to raccoons, their intermediate hosts, and other animals in some areas of the current raccoon range. It should be noted that parasites often move from host to host when the final host consumes the intermediate hosts, but this is not always the case. For example, parasites in genus Schistosoma leave the intermediate host (some aquatic mollusk) and directly penetrate the tissue of the final host (usually primates). Also, data suggest that raccoon roundworm prevalence correlates with behavioral changes in raccoons that make it more likely for the parasites’ eggs to be consumed by intermediate hosts (Ingle 2013). This is accomplished when raccoons select for a diet rich in plant material (the seeds in raccoon feces attract small mammals and other potential hosts). Raccoons from townships with high B. procyonis prevalence ate significantly more plant material than raccoons from townships with low prevalence (χ2 = 7.353, df = 1, p = 0.007; unpublished data). In contrast to B. procyonis, the rat tapeworm, Hymenolepis diminuta, directly interferes with the defense mechanisms of the confused flour beetle (Tribolium confusum) by down-regulating the gene expression for prophenoloxidases, which are involved in the self-recognition and melanization (Hitchen, Shostak, and Belosevic 2009). Raccoon roundworm, the rat tapeworm, and many other highly prevalent parasites provide support for the hypothesis that symbiotic animals were created to make horizontal gene transfer of AGEs possible and efficient.
Finally, symbiotic animals that now live as parasites may have been created to provide some selective pressure on hosts. Thus, these symbionts would have served as a way of bringing the environment inside the dispersing and adapting host. In so doing, symbiotic organisms could have served two major functions: encouraging particular combinations of alleles in offspring and limiting reproduction of species had death not entered into creation with the Fall of Genesis 3. Francis (2009) addressed the function of reproductive control. He postulated that symbiotic organisms might have been created to provide a reproduction control mechanism in a pre-Fall creation. After the Fall brought death into the reality of creation (Genesis 3), these control mechanisms became a parasitic reduction of fitness. Many parasites function to reduce host fitness by restricting or eliminating reproductive output. Francis (2009) noted the impact of the bacterium Wolbachia as it colonizes the reproductive tracts of many insect species, often functioning to reduce reproductive output. Many animal parasites can reduce host reproductive output as well. Schistocephalus causes the gonads of sticklebacks (family Gasterosteidae) to develop for a shorter period of time than the gonads of uninfected sticklebacks (Barber, Hoare, and Krause 2000). The total eggs produced by a pair of T. confusum beetles decreases with increased H. diminuta intensity (Shostak 2009). Female T. confusum beetles have fewer eggs found in the oviduct even with low parasite intensity (Shostak 2012). Because of parasitism like this in animals, the males of hosts subject to high levels of parasitism often show morphological evidence of previous episodes of survived parasitism (Thomas et al. 2000). These parasites may be reflecting the original created design for slowing or eliminating reproductive output once the earth was filled.
In addition to introducing selective pressure in the way of restricting reproduction, parasites introduce selective pressure by encouraging certain combinations of alleles in individual hosts. Parasites exert selective pressure on the host equal with that of predators (Barber, Hoare, and Krause 2000; Thomas et al. 2000). While parasitism and predation were likely not part of the original design for creation, the selective pressure would have been necessary for generating diversification within the created baramins (Wood 2002). Srithayakumar et al. (2011) found a strong correlation between raccoon alleles for Prlo-DRB (a section of the major histocompatibility complex [MHC]) and raccoon rabies virus infection. This correlation demonstrates the selection on certain alleles provided by infectious agents. Similarly, raccoon roundworm functions like many animal parasites by providing selective pressure on hosts, especially in areas with high prevalence (Ingle 2013). Rates of heterozygosity in exon 2 of MHC II differed among townships (Table 1; unpublished data). Heterozygosity ranged from 0.50 in Springfield township to 1.00 in Harmony township. Raccoons from townships with prevalence above 60% had significantly lower heterozygosity than other raccoons from townships with prevalence below 60% (χ2 = 13.13, p < 0.001; unpublished data). Xenia township had the lowest heterozygosity of any township (0.60) with more than five raccoons trapped, and had the highest recorded B. procyonis abundance (18.51 worms per raccoon). These data indicate that the parasite is causing certain alleles to become more common, increasing the number of individuals with two copies of the favored alleles (homozygote) and reducing the number of individuals with two different alleles (heterozygote). Several phylogenetic trees demonstrate evidence for a correlation between raccoon genetic structuring and B. procyonis prevalence. A maximum likelihood (ML) tree shows strong support for grouping Springfield, German, and Greene townships together (Fig. 1; unpublished data). These townships are close in proximity, and all have B. procyonis prevalence below 60%. Maximum likelihood analysis also supports grouping Moorefield and Harmony townships apart from the other seven townships. Harmony has the highest B. procyonis prevalence (73.1%), but Moorefield township has the second lowest recorded prevalence at 46.1%. Both these townships are in Clark County, along with Mad River, Springfield, German, and Greene townships. The neighbor joining (NJ) tree also supports grouping Springfield, German, and Greene townships together, but without as high a bootstrap value as the ML tree (63 and 87, respectively). Harmony and Moorefield townships grouped away from the rest in the NJ tree as well, with Moorefield township grouping closer to the other townships than Harmony (Fig. 2; unpublished data). The three townships surveyed from Greene County, (Beavercreek, Xenia, and Miami townships) grouped along with Mad River township from Clark County, with Beavercreek and Xenia townships form a clade apart from the other two. Beavercreek has the lowest recorded B. procyonis prevalence (24.5%), and grouped with three townships with B. procyonis prevalence above 60%. Finally, the maximum parsimony (MP) tree also shows support (81.5 bootstrap value) for grouping Springfield, German, and Greene townships together (Fig. 3; unpublished data). Maximum parsimony analysis also showed strong support for grouping Moorefield and Harmony townships apart from the other seven. Beavercreek, Xenia, Miami, and Mad River townships also grouped together in the MP tree. Each of the phylogenetic trees was constructed using the sequences from raccoons for the mitochondrial locus, NADH5. The phylogenetic trees from NADH5 also demonstrated evidence of genetic structuring that was more complex than geographic distance alone can explain. Adjacent townships were more likely to group together, although some townships did not group with adjacent townships. Moorefield and Harmony townships always grouped apart from other, even adjacent, townships. Regardless of whether symbiotic animals were created to generate selective pressure on their hosts or not, they continue to provide pressure that makes certain alleles and combinations of alleles more successful than others.
Table 1. Observed heterozygosities for three loci from raccoons from nine townships of southwestern Ohio.
| Township | Number of Raccoons Sampled for MHC II | OH* for MHC II |
|---|---|---|
| Beavercreek | 20 | 0.90 |
| Xenia | 20 | 0.60 |
| Miami | 27 | 0.93 |
| German | 6 | 0.83 |
| Greene | 15 | 0.89 |
| Harmony | 10 | 1.00 |
| Mad River | 8 | 0.75 |
| Moorefield | 12 | 0.83 |
| Springfield | 4 | 0.50 |
| >60%** | 65 | 0.80 |
| <60%*** | 57 | 0.86 |
| * Observed heterozygosity | ||
| ** Townships with B. procyonis prevalence above 60%: Xenia, Miami, Harmony, and Mad River. | ||
| *** Townships with B. procyonis prevalence below 60%: Beavercreek, German, Greene, Moorefield, and Springfield. | ||
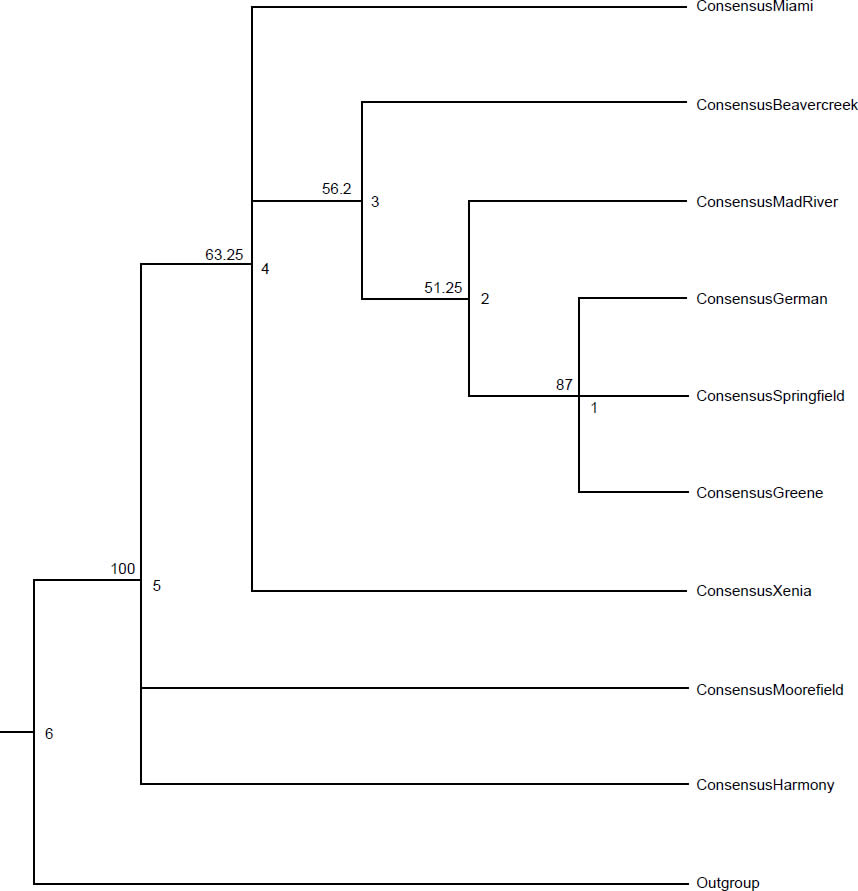
Fig. 1. Maximum likelihood tree for the raccoons from the nine townships of Clark and Greene Counties, Ohio. The branch labels represent the bootstrap support from 1000 pseudoreplicates. The node labels represent the node heights at each node, and the tip labels are the consensus sequences for the corresponding township. We calculated the consensus sequences using a strict method (50% consensus for each base in the sequence). The outgroup is the published sequence for NADH5 from a raccoon from Indiana.
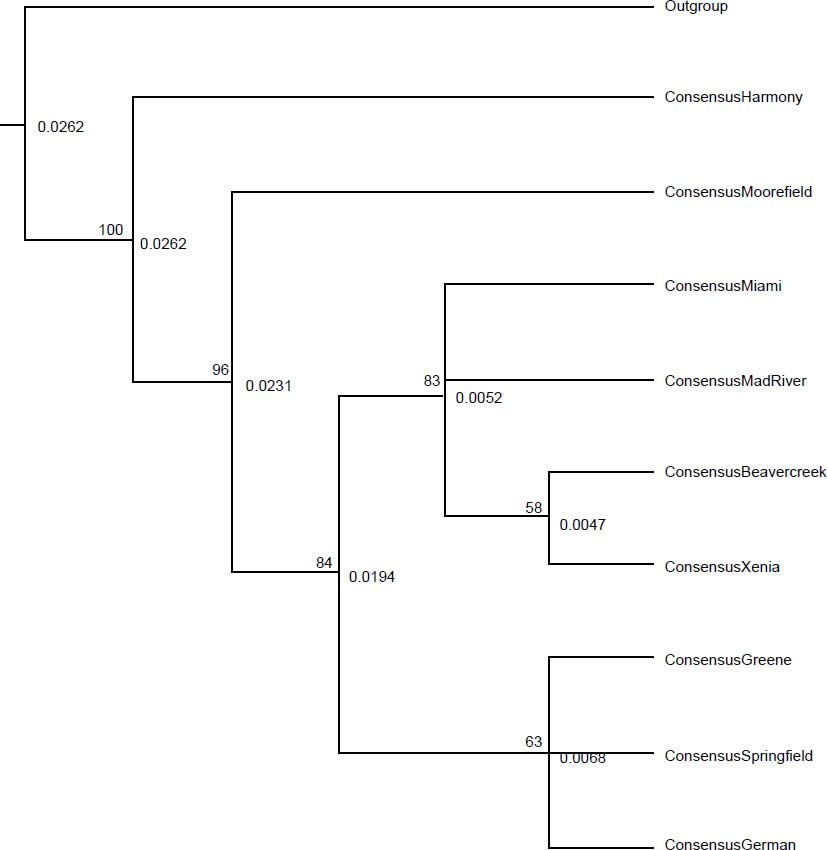
Fig. 2. Neighbor-joining tree for the raccoons from nine townships of Clark and Greene Counties, Ohio. The branch labels represent the bootstrap support from 1000 pseudoreplicates. The node labels represent the node heights at each node, and the tip labels are the consensus sequences for the corresponding township. We calculated the consensus sequences using a strict method (50% consensus for each base in the sequence).
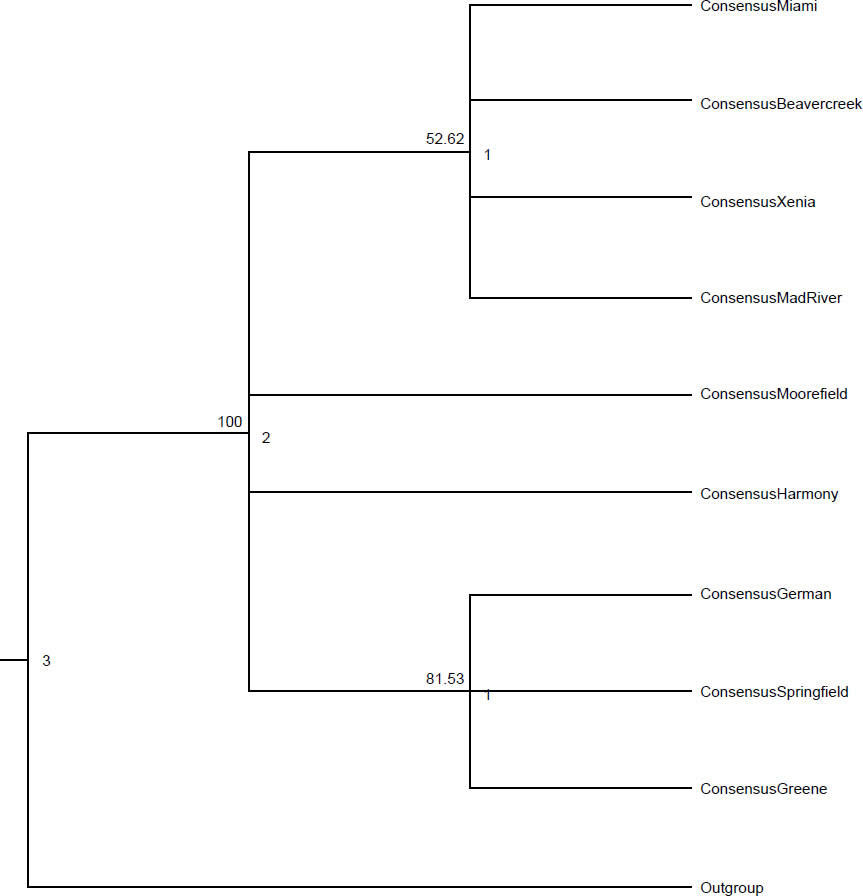
Fig. 3. Maximum parsimony tree for the raccoons from nine townships of Clark and Greene Counties, Ohio. The branch labels represent the bootstrap support from 1000 pseudoreplicates. The node labels represent the node heights at each node, and the tip labels are the consensus sequences for the corresponding township. We calculated the consensus sequences using a strict method (50% consensus for each base in the sequence).
Conclusion
God continues to reveal truths about Himself, His creation and our role in it. As we study biology, other sciences and other disciplines, we learn more about God and how we can serve Him. An honest and fulfilling study of biology requires determining what Scripture says about why and how to study biology, and discovering ways to better steward creation that man has been put in dominion over. As scientists, we are more intimately confronted by God’s creation than anyone else, and this intimacy ought to shape the way we think about God and His creation. God declares His creation “good” throughout Genesis 1. Parasites, reducing the fitness of their hosts, do not fit in this kind of design. Therefore, the organisms that now live as parasites must have been designed to fulfill a function that is either no longer operating or not operating in the same way it did before the Fall in Genesis 3. Three of these functions may have been: functioning as part of the baramin of the host, functioning as vehicles of horizontal gene transfer, and providing the selective pressure necessary for diversification.
References
Barber, I., D. Hoare, and J. Krause. 2000. Effects of parasites on fish behaviour: A review and evolutionary perspective. Reviews in Fish Biology and Fisheries 10:131–165.
Beck, M. A., C. P. Goater, D. D. Colwell, and B. J. van Paridon. 2014. Fluke abundance versus host age for an invasive Trematode (Dicrocoelium dendriticum) of sympatric elk and beef cattle in southeastern Alberta, Canada. International Journal for Parasitology: Parasites and Wildlife 3, no. 3:263–268.
Blank, D., and W. Yang. 2014. Behavioral responses of goitered gazelle (Gazella subgutturosa) to parasitic activity of botflies. Journal of Parasitology 100, no. 1:66–72.
Bloom, P. 2005. Is God an accident? The Atlantic Monthly, 105–112. http://www.theatlantic.com/magazine/ archive/2005/12/is-god-an-accident/304425/.
Boyd, S. W. 2008. The genre of Genesis 1:1–2:3: What means this text? In Coming to grips with Genesis: Biblical authority and the age of the earth, ed. T. Mortenson and T. H. Ury. Green Forest, Arkansas: Master Books.
Butkus, R. A. 2002. The stewardship of creation. The Center for Christian Ethics at Baylor University. http://www.baylor.edu/christianethics/CreationarticleButkus.pdf.
Dawkins, R. 1976. The selfish gene. Oxford, United Kingdom: Oxford University Press.
Dockery, D. S. 2008. Renewing minds: Serving church and society through Christian higher education. Nashville, Tennessee: B&H Publishing Group.
Dunn, R. R., N. C. Harris, R. K. Colwell, L. P. Koh, and N. S. Sodhi. 2009. The sixth mass coextinction: Are most endangered species parasites or mutualists? Proceedings of the Royal Society Biological Sciences 276, no. 1670:3037–3045.
Francis, J. W. 2009. Symbiosis, relationship and the origin of species. In Genesis kinds: Creationism and the origin of species, ed. T. C. Wood and P. A. Garner, Vol. 5. Eugene, Oregon: Wipf and Stock,163–192.
Francis, J. W. 2013. A creationist view of the mammalian immune system: From the Red Queen to social interface. Paper presented at the Origins Conference, August 4, 2013, Pittsburgh, Pennsylvania.
Garner, P. A. 2009. Evolving Christian views of species. In Genesis kinds: Creationism and the origin of species, ed. T. C. Wood and P. A. Garner, Vol. 5. Eugene, Oregon: Wipf and Stock, 7–29.
Gómez, A., and E. Nichols. 2013. Neglected wild life: Parasitic biodiversity as a conservation target. International Journal for Parasitology: Parasites and Wildlife 2:222–227.
Granath, W. O. 2014. Effects of habitat alteration on the epizootiology of Myxobolus cerebralis, the causative agent of salmonid whirling disease. Journal of Parasitology 100, no. 2:157–165.
Hitchen, S. J., A. W Shostak, and M. Belosevic. 2009. Hymenolepis diminuta (Cestoda) induces changes in expression of select genes of Tribolium confusum (Coleoptera). Parasitology Research 105, no. 3:875–879.
Ingle, M. E. 2013. Population genetics of raccoons and raccoon roundworm. Journal of Creation Theology and Science Series B: Life Sciences, Vol. 3.
Ingle, M. E., S. G. Dunbar, M. A. Gathany, M. M. Vasser, J. L. Bartsch, K. R. Guffey, C. J. Knox, A. N. Nolan, C. E. Rowlands, and E. C. Trigg. 2014. Predicting Baylisascaris procyonis roundworm prevalence, presence and abundance in raccoons (Procyon lotor) in southwestern Ohio using landscape features. International Journal for Parasitology: Parasites and Wildlife 3, no. 2:113–117.
Jones, W. I. 2013. “Nature of science.” Unpublished classroom material.
Longman III, T. 2012. Job. Baker commentary on the Old Testament wisdom and Psalms. Grand Rapids, Michigan: Baker Academic.
McElroy, E. J., and de Buron, I. 2014. Host performance as a target of manipulation by parasites: A meta-analysis. Journal of Parasitology 100, no. 4:399–410.
Morris, L. 1976. I believe in revelation. Grand Rapids, Michigan: Wm. B. Eerdman’s Publishing Co.
Anonymous. 2013. Cedarville University doctrinal statement. https://www.cedarville.edu/why-cedarville/doctrinal-statement.
Packer, J. I. 1973. Knowing God. Downer’s Grove, Illinois: Intervarsity Press.
Poulin R., and S. Morand. 2000. The diversity of parasites. The Quarterly Review of Biology 75, no. 3:277-293.
Rossiter, W. 2013. Current opinions: Zeros in host-parasite food webs: Are they real? International Journal for Parasitology: Parasites and Wildlife 2:228–234.
Shostak, A. W. 2009. Tapeworm (Hymenolepis diminuta) infection in flour beetles (Tribolium confusum): Does it cause a trade-off between host fecundity and egg size? Canadian Journal of Zoology 87, no. 11:1087–1095.
Shostak, A. W. 2012. Sequential and concurrent exposure of flour beetles (Tribolium confusum) to tapeworms (Hymenolepis diminuta) and pesticide (diatomaceous earth). Journal of Parasitology 98, no. 3:453–459.
Srithayakumar, V., S. Castillo, R. C. Rosatte, and C. J. Kyle. 2011. MHC class II DRB diversity in raccoons (Procyon lotor) reveals associations with raccoon rabies virus (Lyssavirus). Immunogenetics 63, no. 2:103–113.
Thomas, F., R. Poulin, J.-F. Guegan, Y. Michalakis, and F. Renaud. 2000. Are there pros as well as cons to being parasitized. Parasitology Today 16, no. 2:533–536.
Von Rad, G. 1972. Wisdom in Israel. Trans. J. D. Martin. Harrisburg, Pennsylvania: Trinity Press International.
Wise, K. P. 2013. The illustrations of God’s nature as an explanation of biological phenomena. Paper presented at the Origins Conference, August 4, 2013, Pittsburgh, Pennsylvania.
Wood, T. C. 2002. The AGEing process: Rapid post-Flood intrabaraminic diversification caused by Altruistic Genetic Elements (AGEs). Origins (GRI) 54:5 Wood, T. C., and P. A. Garner. 2009. The real debate over creationism and species. In Genesis kinds: Creationism and the origin of species, T. C. Wood and P. A. Garner, Vol 5. Eugene, Oregon: Wipf & Stock, 1–5.
Zhang, L., D. Hou, X. Chen, D. Li, L. Zhu, Y. Zhang, and J. Li. 2012. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Research 22, no. 1:107–126.









