The views expressed in this paper are those of the writer(s) and are not necessarily those of the ARJ Editor or Answers in Genesis.
Abstract
The anatomy and physiology of temperature regulation was reviewed. The review supports the view that the temperature-regulating systems in animal life are inherently and irreducibly complex. Life can survive only if a relatively narrow temperature window is maintained. For humans, only 7° above normal can be lethal. For some birds and mammals the allowable range is greater, but still within certain limits. Several attempts to explain this complexity by evolution were also reviewed, concluding that evolutionists have not even been able to postulate “just-so” stories to explain their evolutionary theories. They admit that, although temperature regulation is an important consideration in evolution, no progress has been made towards viable explanations of the evolution of these systems in the past century.
Keywords: failure of evolutionary theories, temperature regulation, temperature window, irreducible complexity, cold-blooded animals, warm-blooded animals
Introduction
Heat is lost in all animals by three main means: convection, conduction, and radiation. Conduction includes skin contact with cold water and air, which cools the body. Convection includes cooling by body heat rising. And lastly, all life gives off thermal radiation, specifically infrared radiation (electromagnetic radiation with a frequency lower than visible light). This radiation is perceived by life as heat. The two major thermoregulation designs in vertebrates are the cold-blooded system (such as poikilothermy) and the warm-blooded system (endothermy). In general, all birds and mammals are warm-blooded and all other animals are cold-blooded. This includes not only other vertebrates, such as fish, amphibians, snakes, and most other reptiles, but also, with some exceptions, non-vertebrates such as worms and insects.
The evolution from cold- to warm-blooded systems is considered by evolutionists to be one of the most important evolutionary events in history (Jessen 2012). Evolutionists have also conceded that metabolically based maintenance of “relatively stable body temperature in the face of greatly fluctuating ambient temperature is among the most remarkable attributes of mammals and birds” (Ruben 1995). They believe that this remarkable attribute evolved in mammals about 233 million years ago in the Late Triassic (Araujo et al. 2022; Irving 2022).
The Contrast Between Warm- and Cold-Blooded Animals
Major dissimilarities exist between warm- and cold-blooded animals. So far, attempts to explain the evolution of the two main thermoregulation systems by some theoretical evolutionary scenario have failed. The most authoritative temperature regulation texts do not even attempt to explain the possible evolution of any temperature regulation system (Blatteis 2001; Jessen 2011). The term “evolution” is not in the index of the 294-page book by Blatteis. Attempts to determine when warm-bloodedness in mammals first evolved have also failed.
Temperature Regulation is Critical for Life
Internal temperature regulation is not a minor concern. For humans and most mammals, temperatures below 94° and above 107° Fahrenheit are potentially lethal. The core “body temperature is one of the most tightly regulated parameters of human physiology. At any given time, body temperature differs from the expected value by no more than a few tenths of a degree” (Kurz 2008, 627). Maintaining the proper body temperature is a critical requirement for life’s biochemistry for many reasons. First of all, it is required for the proper functioning of organic catalysts called enzymes. All life-forms use enzymes, which are proteins that regulate chemical reactions. Enzymes are essential for almost all chemical processes in life, including food digestion, liver function, body growth, blood coagulation, wound healing, disease prevention, and even breathing, as well as asexual and sexual reproduction.
If the body temperature is too cold, enzymes work poorly, if at all. If too warm, enzymes denature, losing the physical shape required to function and rendering them nonfunctional. Non-functioning enzymes are disastrous for all cell types. Similarly, either the too cold or too warm problem renders a cell or cellular process inadequate, or it ends the life of the cell. The loss of key enzyme functions is lethal (Osilla, Marsidi, and Sharma 2022).
The Cold-Blooded System
The temperature of cold-blooded animals largely follows changes in their environment, and can be regulated internally only within limits. Many cold-blooded animals regulate their internal body temperature by moving to warmer or cooler environments. Cold-blooded animals cannot remain active in extremely cold temperatures, and in hot, tropical zones, cold-blooded creatures must be careful not to overheat. They can survive by employing certain strategies, such as staying in the shade during the hottest parts of the day.
Cold-blooded animals rely on three basic thermoregulation mechanisms, all requiring different designs. Poikilothermy occurs when the animal’s internal temperature can vary slightly, but its core temperature remains close to the ambient (surrounding) temperature of its immediate environment (Blatteis 2001). One example of poikilothermy is a fish whose gills are highly efficient heat exchangers, allowing it to maintain a consistent core temperature (Nelson, Heath, and Prosser 1984, 793).
Many animals can also utilize external means of regulating body temperature, such as basking in the sunlight, or moving to warmer water (Unger, Rollins, and Thompson 2020, 227). Some crocodiles and other lizards live in the water during the hot season and migrate towards the land during the colder season to burrow into small cavities at the shore area to keep warm (Martin 2017).
Heterothermy describes animals that can self-regulate their body temperature but also, if conditions require it, utilize the surrounding environment to help stay warm or cool. Although some animals exhibit some of the characteristics of both poikilothermy and homeothermy, they do not provide evidence for evolution (Lovegrove 2012).
Regional heterothermy describes organisms that maintain different temperatures in different body regions, usually the limbs. One mechanism to achieve this is countercurrent heat exchangers, such as those used in mammals and birds (Rummel, Swartz, and Marsh 2019). Countercurrent heat exchangers involve blood flowing from the body core to the legs and feet, which carries heat that can be readily lost through the leg or foot skin. In the countercurrent heat exchange design, the vein returning blood to the body core lies alongside the artery taking blood to the feet. Heat then moves by conduction from the warmer arterial blood to the cooler venous blood, reducing heat loss from the body.
Body temperature changes necessitate that the animal move to an environment that can maintain its required body temperature range. This requires the ability to sense the temperature of the surrounding environment. If the temperature is not at the necessary level for the animal to maintain its required body temperature, a move to somewhere else is mandatory. Achieving this awareness requires a sensory system and a brain that can coordinate the animal’s proper response to external temperature requirements. Most cold-blooded animals utilize a combination of the above thermoregulation mechanisms to achieve this goal (Blatteis 2001).
Problems With the Cold-Blooded to Warm-Blooded Evolution Theory
Although evolutionists commonly assumed that warm-blooded animals evolved from their cold-blooded relatives, some poikilotherms have more complex metabolisms than their warm-blooded counterparts. For example, in order to thrive in different temperatures certain poikilotherms require up to ten different enzyme systems, instead of one (Blatteis 2001). In this case, poikilothermy is genetically more complex than that of warm-blooded animals occupying the same ecological niche. The reason is that each of the ten enzyme systems requires a different set of genes to produce and regulate it compared to the one required for a single enzyme system. Thus, the simplistic cold-blooded to warm-blooded evolutionary scenario is problematic from step one.
Temperature Regulation in Endotherms
Warm-blooded animals are capable of maintaining a nearly constant internal body temperature, usually close to 35–40°C, irrespective of their environmental temperature (Polymeropoulos, Oelkrug, and Jastroch 2018, 891). Their body temperature remains stable even when the animal moves to another environment with a different average temperature. To achieve this temperature consistency, they use a set of complex internal mechanisms including metabolic regulation that helps them stay warm in colder areas, and cool in warmer areas.
Homeothermic species maintain a stable body temperature by regulating metabolic processes. Critical in achieving this internal temperature consistency is the preoptic area (POA) of the hypothalamus. The POA is located in the brain stem and connected to both the cerebrum and the spinal cord. Its main function is to keep the body in a stable temperature state called homeostasis by directly influencing the autonomic nervous system, or by managing hormone balance and levels.
Endothermy warms tissue by the production of metabolic heat, such as by burning fat (Clarke 2017). Homeothermy thermoregulation uses physiological means such as sweating and shivering instead of behavioral means such as moving to another location (Akin 2011). Sweating and shivering are both complex systems controlled by the hypothalamus and the nervous/hormonal system (Kurz 2008). Shivering is triggered only when the body insulation and environment is insufficient to maintain the required body temperature. It involves rapid muscle contractions that use ATP, consequently producing heat.
Another way that mammals maintain a constant internal temperature when living in varying external temperatures is by growing thick fur during winter to help conserve body heat and shedding it during summer to facilitate heat loss. This method is commonly used by furry mammals, including horses, bison, wolverines, mountain goats, sheep, and some dogs. Feathers used by birds serve the same function. Mammals also typically have well-developed insulation to retain body heat—blubber in the case of marine mammals (Blatteis 2001).
A brief review of this system illustrates the complexity of thermoregulation. Animals can shunt their blood to or away from the skin to radiate heat for cooling off, which requires coordination of the nervous system-controlled cardiovascular system. They also use behavioral strategies to cool themselves off by convection, conduction, or radiation. Some will go for a swim, stand under trees, wallow in mudholes, sleep in the hottest part of the day and come out only at night, or use other means to cool their body. During winter, some mammals maintain metabolic activities during hypothermia via a process called hibernation. Hibernation involves slowing down metabolism in order to spend the winter in a state of dormancy, using long-term, multi-day torpor for survival during cold conditions. Animals in hot climates also undergo a form of hibernation called estivation that enables them to survive extreme heat, drought, or lack of food. Estivation is the use of dormancy during the summer to survive hot and dry periods (Storey and Storey 2010).
Sweat Glands
No cold-blooded animal has sweat glands; only warm-blooded animals do. Humans have two types of sweat glands—eccrine and apocrine glands (fig. 1). In total, we have between two and five million sweat glands located throughout our body. All other primates and mammals have only apocrine glands. With few exceptions, mammals are covered with thick hair, consequently they must have localized sweat glands on their paws. This includes horses, monkeys, apes, dogs, and mice. In hot environments, water evaporation cools by shedding excess heat, either by sweating in many mammals, or by short quick breaths called panting. Many mammals and all birds pant. Cool air from panting comes into contact with the moist lung lining and throat, lowering body temperature. The cool air also causes water evaporation from the tongue and throat as well as the lung lining, helping to further cool the body.
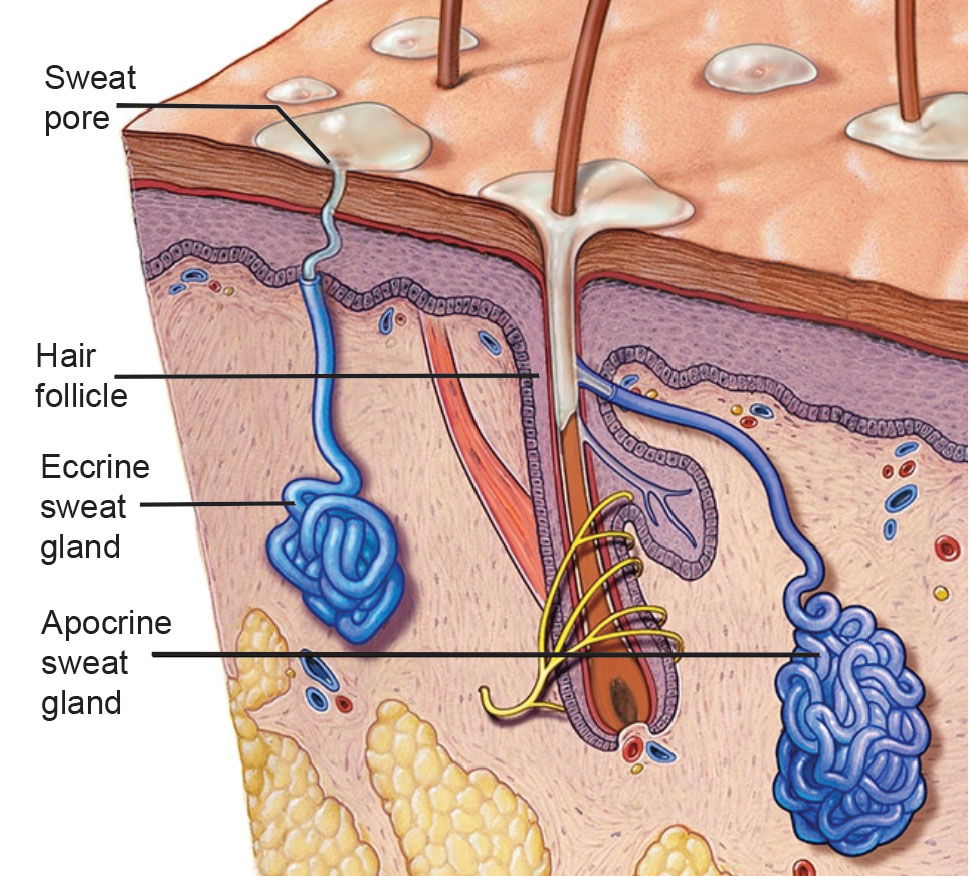
Fig. 1. An illustration of the contrast between Eccrine and Apocrine sweat glands (from the Mayo Clinic Education Foundation.)
Of special note is that vigorous deep-breathing by humans can cause problems in maintaining body acid-base balance, arterial diameter, and sodium retention by the kidneys. Animals that use panting do not have this problem because they are designed to employ very shallow pants. They have mechanisms to determine the appropriate frequency and depth of panting to avoid problems, which illustrates design by our Creator.
Body heat is generated by metabolism via chemical reactions in cells that break down glucose into water and carbon dioxide. This reaction generates ATP (adenosine triphosphate), a high-energy compound required to power most cellular processes. Around 60% of the available energy from metabolism is converted into heat rather than to ATP. In cold-blooded organisms, this heat is lost to the environment. Conversely, warm-blooded endothermic homeotherms produce more heat than is required for normal body functions. They then release the excess heat to help regulate body temperature.
Evolution of Thermoregulation
In an extensive literature review I was unable to find any viable evidence for the actual physical evolution of ectotherms into endotherms. Even references titled “the evolution of thermoregulation” lacked “just-so” stories of plausible mechanisms. The problem for evolution is that most thermoregulation mechanisms require a complex network of special receptor, both internally and externally, plus nerves, muscles, respiratory, circulatory and hormonal control sites, including organs such as the heart, the brain, and the preoptic area (the anterior part of the hypothalamus). One recent literature review of the evolution of thermoregulation mechanisms concluded that the
independent acquisition of endothermy by mammals and birds has been the subject of many hypotheses regarding their origin and associated evolutionary constraints. Many groups of vertebrates, however, are thought to possess other mechanisms of heat production, and alternative ways to regulate thermogenesis that are not always considered in the paleontological literature. . . . endothermy in mammals and birds is not as well defined as commonly assumed by evolutionary biologists and consists of a vast array of physiological strategies, many of which are currently unknown. (Legendre and Davesne 2020)
The problem for evolution is irreducible complexity, as explained by Polymeropoulos, Oelkrug, and Jastroch as follows: “Metabolic heat production required a complex degree of coordination from the organism . . . to achieve a high, stable body temperature (Tb), or homeothermic endothermy . . . . a key trait of birds and mammals” (Polymeropoulos, Oelkrug, and Jastroch 2018, 981). The supposed evolutionary benefits are described as “resulting expansion of endotherms into thermal niches that were not accessible to ectotherms, whose Tb is dictated by ambient temperature (Ta), proved to be the crucial evolutionary advantage” (Polymeropoulos, Oelkrug, and Jastroch 2018, 981). The challenge of thermoregulation evolution was outlined by Bakker as follows:
Among living tetrapods the most striking contrast in bioenergetics is that between ectotherms, represented by lizards, and endotherms, represented by birds and advanced placental mammals. Since both birds and mammals evolved from ectothermic reptiles, consideration of this contrast is a useful starting point for reviewing the fossil record. (Bakker 1975)
Evolutionists’ problem is that, “the evolutionary framework and events leading to endothermy are still unclear . . . . Tracing back the heterothermic ancestor giving rise to strict homeothermy has been a matter of a vivid debate” (Polymeropoulos, Oelkrug, and Jastroch 2018, 981). Polymeropoulos, Oelkrug, and Jastroch then state that “three streams of hypotheses have been proposed aiming to explain this extraordinary transition from ecto- to endothermy during vertebrate evolution.” These hypotheses are not steps that explain how they evolved but why they evolved, which translates into the advantages that resulted from their evolution. The three hypotheses, actually guesses, are as follows:
The first school of thought promotes that the thermoregulatory advantages of high Tbs per se, resulting in higher metabolic rates (MR), suffice to explain the evolutionary advantage. Others have put forward [the idea] that the selection for higher MRs is a consequence of increased exercise, consequently raising Tb. More recently, the parental care hypothesis has gained more traction, which stipulates an increase in Tb being beneficial for growth while reducing mortality, thus increasing species’ fitness. (Polymeropoulos, Oelkrug, and Jastroch 2018, 981)
Another example illustrated the “suggestion” level of science. Italics were added to emphasize the tentative nature of the “suggestions” which follow:
It is suggested that small endotherms cannot be directly derived from small ectotherms because of the requirement for the simultaneous change in thermal conductance and the rate of metabolism. Instead, small ectotherms probably gave rise to large ectotherms (thereby increasing thermal inertia), a fur coat may have been added to increase thermal stability, and a decrease in size with only a modest decrease in the total rate of metabolism converted “inertial” homoiothermy to endothermy . . . . A decrease in size and the formation of a secondary palate also occurred in bauriamorphs, which suggests that they too independently evolved endothermy, although it may not have been as effective as that developed in cynodonts. (McNab 1978)
In articles dealing with the evolution of thermoregulation, terms such as development or acquisition were often used instead of evolution (Nelson, Heath, and Prosser 1984). Nelson, Heath, and Prosser used development 16 times, acquisition three times. Terms that are commonly used to indicate speculation, not evidence, were often noted, such as may have, used by Nelson 26 times, likely, used six times, and probably, used 13 times. Thus, in a 14-page article, terms that are commonly used to indicate speculation were used a total of 45 times, or 3.2 times per page. Satinoff (1978), quoted in Nelson, postulated that “nearly all thermoregulation effectors of tetrapods, including mammals, were appropriated from pre-existing voluntary and autonomic patterns” (Nelson, Heath, and Prosser 1984, 802). Furthermore, evolution is proposed to have occurred to “solve specific problems,” an explanation reminiscent of Lamarckism:
The orchestration of the responses of a series of semi-autonomous subsystems, each with thermosensitivity and probably each appropriating and elaborating pre-existing motor patterns during evolution to solve specific adaptation problems, forms a special challenge to future adaptational biologists. (Nelson, Heath, and Prosser 1984, 805)
Claiming that thermogenesis evolved, but not theorizing how it could have evolved, is the norm in the journals since Nelson published his review in 1984. Typical is an article regarding the origin of shivering thermogenesis which stated only that it evolved, but nothing about how it could have evolved (Haman and Blondin 2017, 217). As irreducibly complex systems which require the numerous subsystems outlined above, the thermoregulation systems could not function to regulate the temperature until all of the required components were in place and functioning as a unit. This fact was acknowledged by Nelson, but he did not even attempt to explain the origin of the systems:
How did the complexly controlled multiple effector system come about? Central mechanisms for thermoregulation apparently did not evolve de novo, but rather in connection with other functions. Regulatory changes which appeared were more likely quantitative rather than qualitative. (Nelson, Heath, and Prosser 1984, 802)
How this gradual evolution could have been produced by evolution was not detailed. Satinoff (1978) argued that current theories of thermoregulation were wrong. He then speculated that “several hierarchically arranged integrating systems may have evolved to achieve precise thermoregulation.” The progress in explaining the small steps of thermoregulation evolution is today close to where it was almost a half-century ago.
For example, Angilletta et al., when proposing new insight into the evolution of thermoregulation, admitted that “the dearth of empirical data on this subject [of the evolution of thermal physiology] precludes a rigorous analysis at this time” (Angilletta et al. 2010). They thus “urge researchers to investigate the evolution of thermal physiology in endotherms . . . . [to] generate new lines of inquiry about the evolution of their thermal physiology” (Angilletta et al. 2010, 862, 864). Furthermore, they ask the following question which they never attempt to answer:
what macroevolutionary patterns should we expect to observe among species of mammals and birds? Although we cannot answer these questions at present, we speculate about the answers in the sections that follow . . . . The evolution of thermosensitivity is further complicated by the fact that some body tissues are warmer than others. (Angilletta et al. 2010, 871–873)
Although “regional and temporal heterothermy in endotherms should drive the evolution of thermosensitivity, at present, we know virtually nothing about the thermosensitivity of performance in mammals and birds . . . . [and basic] questions regarding the evolution of thermal physiology remain entirely unresolved” (Angilletta et al. 2010, 874–875).
Numerous papers were located that attempted to explain why the putative evolution from cold-bloodedness to warm-bloodedness occurred, as well as its importance, but ignore the question how it did, or even how it could have evolved in the small steps Darwinian evolution requires (Dawson 1972). Nonetheless, evolution was assumed to be the correct explanation for the origin of each system described. Typical is the following observation: “an increase in the number of thermoregulatory effector mechanisms has occurred with evolution in several vertebrate lines” (Nelson, Heath, and Prosser 1984, 793). The evolution of the individual systems required for endothermy was never explained. Another problem for evolution is that some reptiles are believed to have been homeotherms, including ichthyosaurs, pterosaurs, plesiosaurs, and some non-avian dinosaurs. Both systems appear very early in history. Thus the overlap of the two systems, cold-blooded and warm-blooded very early in history, argues against one system evolving into the other system early in evolutionary history as Darwinism has proposed.
Controversy Over When Endothermy Appeared
Although “Endothermy underpins the ecological dominance of mammals and birds in diverse environmental settings . . . . it is unclear when this crucial feature emerged during mammalian evolutionary history, as most fossil evidence is ambiguous” (Araujo et al. 2022, 726). The evolution from cold-blooded into warm-blooded systems requires significant modifications in both anatomy and physiology, modifications that are some “of the most important in evolution” (White 1891, 374). Over a century later no progress has been made to decipher this assumed mystery. One attempt to determine when it occurred proposed a three-phase model in which the first step occurred in response to improved parental care and the demands of conquering dry land. Lovegrove then proposed
that Phase Two commenced in the Late Triassic and Jurassic and was marked by extreme body-size miniaturization, the evolution of enhanced body insulation (fur and feathers), increased brain size, thermoregulatory control, and increased ecomorphological diversity. I suggest that Phase Three occurred during the Cretaceous and Cenozoic and involved endothermic pulses associated with the evolution of muscle-powered flapping flight in birds, terrestrial cursoriality in mammals, and climate adaptation in response to Late Cenozoic cooling in both birds and mammals. (Lovegrove 2017)
This does not explain how these developments occurred by gradual evolution. Darwinian evolution predicts that each anatomical and physiological step must have provided a selective advantage. Darwin’s theory is that evolutionary change or divergence accumulates steadily and slowly, in small steps, as one species succeeds the previous one in the struggle for existence. In his words “natural selection acts solely by accumulating slight, successive, favorable variations, it can produce no great or sudden modifications; it can act only by short and slow steps” (Darwin 1859, 471). Theories of endothermy evolution ignore the slow gradual steps leading up to the functional organ because researchers are not even able to imagine a possible plausible Darwinian scenario. Most academic papers on the evolution of the thermoregulatory system assume evolution and explain theoretically how it could have worked, but not how warm-bloodedness could have evolved. Here is an example:
[one] underlying principle may be unique to the system: thermoregulatory mechanisms are often derived from other organ systems, e.g., the cardiovascular system during vasomotion or the respiratory system in panting. The thermoregulatory system has not evolved specific mechanisms to control body temperature but makes use of pre-existing systems . . . . Thus, an increase in the number of thermoregulatory effector mechanisms has occurred with evolution in several vertebrate lines. (Nelson, Heath, and Prosser 1984, 793)
The problem with this explanation is: Which system evolved first? Did one evolve, then the other? Or did both develop concurrently, simultaneously evolving? The problem is that the animal cannot survive at any point in its development without working, highly functional, thermoregulatory, and cardiovascular systems.
The main theory for why temperature control evolution occurred is over a century old and is still the main theory accepted today. It explains that, in “vertebrates, at least, cold-blooded animals are aquatic and warm-blooded animals are primarily terrestrial” (White 1891, 374). One reason for this general difference is that ocean and lake water temperatures are very consistent from hour to hour. In contrast, land temperature can change as much as 30° Fahrenheit within a few hours.
This ocean/land temperature difference explains why very different thermoregulation designs were required for each animal type in order to survive in its specific environment. Again, it does not explain how the warm-blooded trait could have evolved. A better explanation is that warm-blooded vertebrates were created to live in a terrestrial world, and many cold-blooded vertebrates were created to live in a water world as part of their original design.
Professor White added that, as evolution postulates, life first evolved in water, therefore was originally cold-blooded. For life to live on land, the warm-blooded design was required, but until this system was functional, terrestrial life would be impossible (White 1891, 374). He then explained away the problem of the evolution from water to land by postulating enormous amounts of time to allow for warm-bloodedness to evolve.
The attempted explanation is that the cold-blooded temperature regulation system sufficed on land until life gradually evolved the warm-blooded system. The main attempt to support evolution for the past half century has been to show that the more warm-blooded the animal is, the higher it is on the evolutionary scale; reptiles are the least warm-blooded, man the most (Cloudsley-Thompson 1968). The problem with this reasoning is that a sharp dichotomy exists between cold- and warm-blooded systems, not a gradual progression as evolution requires. Furthermore, evolutionists believe that birds evolved warm-bloodedness independently from mammals, meaning that the warm-blooded system independently evolved at least twice. This theory is an idea that lacks fossil and any other direct evidence.
One article which claimed that scientists have pinpointed “the exact moment” in evolutionary time when mammals became warm-blooded is irresponsible hype (Turner 2022). Other published reports on this research were more cautious, one stating that “Warm-Bloodedness in Mammals May Have Arisen in Late Triassic” (Carstens 2022; italics added). The actual research published in Nature was far less confident about the results, stating that their research only “suggests that endothermy evolved abruptly during the Late Jurassic” (Araujo et al. 2022, 726). The review added that the new research’s conclusion was that “it happened much more quickly than scientists expected.”
Summary
This review of the contrast between warm- and cold-bloodedness shows the lack of a plausible scenario to explain how warm-bloodedness could have evolved from the very different cold-bloodedness design. The chasm between the cold-blooded system and the warm-blooded system is enormous and has not been bridged by evolutionists, even by “just-so” stories. Evolutionists not only have failed to explain how the small steps of evolution from the cold-blooded to the warm-blooded system of temperature regulation could have existed, but also the evolution of other, often interrelated, systems, including poikilothermy, heterothermy, ectothermy, endothermy, and homeothermy. Evolutionists also need to explain the origin of the functionally integrated temperature control system, including the many specialized sensory cells related to temperature control.
Part of the reason for the difficulty of postulating the evolution of the different thermoregulation systems is because all known temperature regulation systems are part of the body’s complex nervous, hormonal, hypothalamic, muscular, glandular, venous, and arterial systems. All these systems must be functionally integrated to maintain the body temperature above the narrow hypothermal lower limit, and below the hyperthermal upper limit. And until the entire thermoregulation system is functional to effectively maintain this narrow range, eukaryotic life would not be possible.
References
Akin, Jonathan A. 2011. “Homeostatic Processes for Thermoregulation.” Nature Education Knowledge 3, no. 10: 7.
Angilletta, Michael J. Jr., Brandon S. Cooper, Matthew S. Schuler, and Justin G. Boyles. 2010. “The Evolution of Thermal Physiology in Endotherms.” Frontiers in Bioscience E2, 2, no. 3 (June 1): 861–881.https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.457.9952&rep=rep1&type=pdf.
Araujo, Ricardo, Romain David, Julient Benoit, Lungmus Jacqueline, Fred Spoor, Alexander Stoessel, Paul M. Barrett, et al. 2022. “Inner Ear Biomechanics Reveals a Late Triassic Origin for Mammalian Endothermy.” Nature 607, no. 7920 (July): 726–731.
Bakker, Robert T. 1975. “Experimental and Fossil Evidence for the Evolution of Tetrapod Bioenergetics.” In Perspectives of Biophysical Ecology, vol. 12, edited by David M. Gates and Rudolf B. Schmerl, 365–399. New York, New York: Springer.
Blatteis, Clark. ed. 2001. Physiology and Pathophysiology of Temperature Regulation. 5 Toh Tuck Link, Singapore: World Scientific Publishing Co.
Carstens, Andy. 2022. “Warm-Bloodedness in Mammals May Have Arisen in Late Triassic.” The Scientist, July 21. https://www.the-scientist.com/news-opinion/warm-bloodedness-in-mammals-may-have-arisen-in-late-triassic-70253.
Clarke, Andrew. 2017. Principles of Thermal Ecology: Temperature, Energy, and Life. New York, New York: Oxford University Press.
Cloudsley-Thompson, John L. 1968. “Hot Blood or Cold? Thermoregulation in Terrestrial Poikilotherms.” Science Progress 56, no. 224 (October): 499–509.
Darwin Charles. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. London, United Kingdom: John Murray.
Dawson, Terence John. 1972. “Primitive Mammals and Patterns in the Evolution of Thermoregulation.” In Essays on Temperature Regulation, edited by J. Bligh and R. Moore, 1–18. Amsterdam, Netherlands: North- Holland.
Haman, François, and Denis Blondin. 2017. “Shivering Thermogenesis in Humans: Origin, Contribution and Metabolic Requirement.” Temperature: Multidisciplinary Biomedical Journal 4, no. 3 (May 22): 217–226.
Irving, Katherine. 2022. “Our Ancestors Became Warm-Blooded Later Than We Thought.” Science, July 20. https://www.science.org/content/article/our-ancestors-became-warm-blooded-later-we-thought.
Jessen, Claus. 2012. Temperature Regulation in Humans and Other Mammals. New York, New York: Springer.
Kurz, Andrea. 2008. “Physiology of Thermoregulation.” Best Practice and Research Clinical Anaesthesiology 22, no. 4 (December): 627–644.
Legendre, Lucas L., and Donald Davesne. 2020. “The Evolution of Mechanisms Involved in Vertebrate Endothermy.” Philosophical Transactions of the Royal Society B 375 no. 1793 (March 2): 20190136.
Lovegrove, Barry G. 2012. “A Single Origin of Heterothermy in Mammals.” In Living in a Seasonal World: Thermoregulatory and Metabolic Adaptations, edited by Thomas Ruf, Claudia Bieber, Walter Arnold, and Eva Millesi, 3–11. New York, New York: Springer Nature.
Lovegrove, Barry G. 2017. “A Phenology of the Evolution of Endothermy in Birds and Mammals.” Biological Reviews of the Cambridge Philosophical Society 92, no. 2 (May): 1213–1240.
Martin, Anthony J. 2017. “The Evolutionary Advantage of Burrowing Underground.” American Scientist 105, no. 5 (January): 306–312.
McNab, Brian K. 1978. “The Evolution of Endothermy in the Phylogeny of Mammals.” The American Naturalist 112, no. 983 (January–February): 1–21.
Nelson, Douglas O., James E. Heath, and C. Ladd Prosser. 1984. “Evolution of Temperature Regulatory Mechanisms.” American Zoologist 24, no. 3 (August): 791–807.
Osilla, Eva V., Jennifer L. Marsidi, and Sandeep Sharma. 2022. Physiology, Temperature Regulation. Treasure Island, Florida: StatPearls Publishing.
Polymeropoulos, Elias T., Rebecca Oelkrug, and Martin Jastroch. 2018. “Editorial: The Evolution of Endothermy—From Patterns to Mechanisms.” Frontiers in Physiology 9 (12 July): 891.
Ruben, John. 1995. “The Evolution of Endothermy in Mammals and Birds: From Physiology to Fossils.” Annual Review of Physiology 57: 69–95.
Rummel, Andrea D., Sharon M. Swartz, and Richard L. Marsh. 2019. “Warm Bodies, Cool Wings: Regional Heterothermy in Flying Bats.” Biology Letters 15, no. 9 (September): 1–5. https://royalsocietypublishing.org/doi/epdf/10.1098/rsbl.2019.0530.
Satinoff, E. 1978. “Neural Organization and Evolution of Thermal Regulation in Mammals: Several Hierarchically Arranged Integrating Systems May Have Evolved to Achieve Precise Thermoregulation.” Science 201, no. 4350 (7 July): 16–22.
Storey, Kenneth, and Janet Storey. 2010. “Metabolic Regulation and Gene Expression During Aestivation.” In Aestivation: Molecular and Physiological Aspects, edited by Carlos Arturo Navas and José Eduardo Carvalho, 25–46. New York, New York: Springer.
Turner, Ben. 2022. “Scientists Pinpoint the Exact Moment in Evolutionary Time When Mammals Became Warm-Blooded.” Live Science, July 21. https://www.livescience.com/warm-blooded-mammals-evolution.
Unger, Shem D., Mark A. Rollins, and Christy M. Thompson. 2020, “Hot- or Cold-Blooded? A Laboratory Activity That Uses Accessible Technology to Investigate Thermoregulation in Animals.” The American Biology Teacher 82, no. 4 (April): 227–233.
White, William Hale. 1891. “A Theory to Explain the Evolution of Warm-Blooded Vertebrates.” Journal of Anatomy and Physiology 25, no. 3 (April 1): 374–385.