

Research conducted by Answers in Genesis staff scientists or sponsored by Answers in Genesis is funded solely by supporters’ donations.
Abstract
The genus Microraptor is classified within the Dromaeosauridae. It was first found in the Early Cretaceous Jiufotang Formation in Liaoning, China. It has played a central role in the debate over the supposed evolutionary relationship between dinosaurs and birds and the supposed “feathered” dinosaurs. This genus has been considered a feathered dinosaur by most secular scientists and some creation scientists. Microraptor has been used to support the hypothesis that birds evolved from dinosaurs and that dinosaurs had feathers. This paper examines the anatomical skull features of the first three described Microraptor species (Microraptor zhaoianus, Microraptor gui, and Microraptor hanqingi) to understand whether they align more closely with bird or dinosaur characteristics. This analysis is performed through a literature review of the first original publications of the first three species of Microraptor. The original descriptions of the bone structures in the publications were compared to determine whether they follow a dinosaurian or bird-like pattern. That was done by comparing the secular literature on extinct birds and dinosaurs.
The findings indicate that the Microraptor’s skull exhibits numerous characteristics that are also shared with extinct birds. This study raises questions about the interpretation of Microraptor as a feathered dinosaur and instead supports its classification as an extinct bird. The result of this analysis can contribute to the ongoing debate about whether those anatomical structures support a dinosaurian or a bird classification.
Keywords: Microraptor, Dromaeosauridae, feathered dinosaurs, dinosaur-bird relationship, cranial morphology, avian characteristics, extinct birds
Introduction
Microraptor is a recognized and classified genus of Dromaeosauridae from the Early Cretaceous of the Jiufotang Formation in Liaoning, Northeastern China. This formation records many specimens of birds and supposedly feathered dinosaurs. This formation is above the Yixian Formation. Both are part of the Jehol Group, which gained fame in the 1990s with the discovery of Sinosauropteryx, the first supposedly feathered dinosaurs.
Microraptor has been accepted as one of the few dinosaur genera identified as having feathers by secular and some creation scientists (Wood and Garner n.d.). Microraptor has played a central role in the ongoing debate about feathered dinosaurs. However, many secular researchers have supported the possibility that the supposedly feathered dinosaurs are birds (Bechly 2024 a, b).
Here, I analyze the morphological features of the first three species of Microraptor that have been identified, described, and published. Since hundreds of specimens represent Microraptor (Alexander et al. 2010), it is impractical to analyze each one of the specimens individually. This analysis focuses on the cranial elements to examine whether they align with a bird or dinosaur structure based on current secular literature.
For now, the scientific consensus is that this genus has three species: Microraptor zhaoianus gen. et sp. nov. (Xu, Zhou, and Wang 2000), Microraptor gui sp. nov. (Xu et al. 2003), and Microraptor hanqingi (Gong et al. 2012). The synonymy (when two or more different scientific names are used for the same fossil species or genus) proposed by Turner, Makovicky, and Norell (2012) and Senter et al. (2004) for this genus is not discussed here.
Based on the anatomical analyses of the skulls of these three species, the view of Microraptor as a feathered dinosaur is contested. The analyzed skull structure was listed, mentioned, identified, and described in the first three original publications of Microraptor specimens (Gong et al. 2012; Xu et al. 2003; Xu, Zhou, and Wang 2000).
Understanding Microraptor as a bird can help support the idea that other specimens with similar features can be classified as birds instead of feathered dinosaurs. That is why evaluating whether the features listed, mentioned, identified, and described for this genus align more with those of birds or dinosaurs is necessary.
The Three Species of Microraptor—General View
Microraptor zhaoianus was discovered with a partially articulated skeleton, lacking the middle portion of the body. One specimen (V 12330) was used for its description (Xu, Zhou, and Wang 2000).
Most of the six specimens found of Microraptor gui were articulated well, and the holotype is an almost complete skeleton. The holotype (IVPP V13352) and a referred specimen were used for its description (Xu et al. 2003).
M. hanqingi is a nearly complete specimen, showing a mostly articulated postcranial skeleton with a disarticulated pectoral girdle and left forearm. One specimen (LVH 0026) was used for its description (Gong et al. 2012).
Nothing is said in the publication about the quality of preservation of the skeletal material for M. zhaoianus and M. gui. Regarding M. hanqingi, it is mentioned that one of the sides of the skeleton exhibits, “well-preserved skeletal elements and details not evident on the original surface” (Gong et al. 2012).
The first specimens analyzed for the publication of M. zhaoianus were identified as mature (Xu, Zhou, and Wang 2000). There is no mention of the degree of maturity of M. gui and M. hanqingi (Gong et al. 2012; Xu et al. 2003).
Skull Bones
In the secular scientific field, information in the skull has been necessary to understanding dinosaur-bird evolution. According to Bhullar et al. (2012), “The unique bird skull houses two highly specialized systems: the sophisticated visual and neuromuscular coordination system allows flight coordination and exploitation of diverse visual landscapes, and the astonishing variations of the beak enable a wide range of avian lifestyles.” Similarly, Felice et al. (2020) highlight that some defining avian traits are localized specifically in the skull, “key ‘avian’ features being localised to the skull (e.g., edentulous beak, kinetic palate, encephalised brain).”
These morphological variations in birds’ skulls emphasized by Bhullar et al. (2012) and Felice et al. (2020), are understood in the secular worldview as results of evolutionary processes. Recognizing that secular scientists interpret anatomical structures through the lens of evolutionary ideas is essential when analyzing any morphological feature in this context. Neutrality does not exist, and the evolutionary worldview has been applied to interpreting traits to tell an evolutionary story of those traits, not accounting for shared design and functionality.
The Three Species of Microraptor and Their Skulls
Not many of the skull features of Microraptor zhaoianus are described and discussed in the original publication (Xu, Zhou, and Wang 2000) of this species. The information mentioned is that there are similarities between specific features of the premaxilla and those of Archaeopteryx and Sinornithosaurus. Additionally, certain features of the maxilla are like those of Archaeopteryx and troodontids (one of the groups that belong to Maniraptora).
On M. gui not much was said in its original publication (Xu et al. 2003) about skull morphology. The morphological information described for the skull included an identifiable triradiate postorbital, one of the bones that contribute to the orbital structure.
The skull of M. hanqingi is the most fully described. The cranial elements are the skull and mandible. They were found to be closely associated. The skull varies in preservation, and many cranial bones can be identified. They are mostly disarticulated. Surface ornamentation is not present. The skull has a length of 92.5 mm, but this measurement is based on the length of the mandible. The identified cranial bones are “right and left maxilla, the remnants of both premaxillae, nasals, lacrimal, squamosal, dentaries, posterior mandibular elements including the articular, surangular, prearticular, angular, and splenial” (Gong et al. 2012).
Premaxilla
A premaxilla is mentioned in the original publications of M. zhaoianus as, “The premaxilla is similar to that of Archaeopteryx and Sinornithosaurus in that it has a sloping anterior margin. As in Archaeopteryx and troodontids, the maxilla contributes to the border of the external naris” (Xu, Zhou, and Wang 2000). And for M. hanqingi as, “Preservation of the skull varies, and the cranial bones that can be identified include the right and left maxilla, remnants of both premaxillae . . .” (Gong et al. 2012)
M. zhaoianus has a sloping anterior margin. This feature in M. hanqingi is indicated based on the outline of the maxilla. Xu and Wu (2001) cited the premaxilla’s sloping anterior margin as a bird-like feature. No information on this structure is noted for M.gui in its original publication.
Maxilla
Not much is mentioned about the maxilla of M. zhaoianus. There is only a comparison of its contribution to the external border of the naris as in Archaeopteryx and troodontids (Xu, Zhou, and Wang 2000). Nothing is said about the maxilla of M. gui.
M. hanqingi has a maxilla that is approximately 43.5 mm long and a height projection of 20.6 mm (Gong et al. 2012). Due to its outline, the skull exhibits a trait like that of Archaeopteryx and Sinornithosaurus. M. hanqingi’s maxilla is preserved with four teeth in the original position. However, there are 14 alveoli in it. The maxillary presents teeth that are longer and more recurved compared to the dentary teeth (Gong et al. 2012).
Dentary
The dentary of M. hanqingi presents an arrangement of alveoli that are close together (Gong et al. 2012). It accommodates as many as 16 teeth, possibly. Therefore, the number of teeth for M. hanqingi is unclear. The dentary has short, broad teeth compared to the maxillary teeth. They also present a uniform size.
M. zhaoianus shows a dentary with at least 15 dentary teeth (Xu, Zhou, and Wang 2000). Its dentary teeth are packed as in troodontids. Troodontids are recognized as a bird group (Feduccia 2020). No information about the dentary of M. gui is mentioned.
Teeth
Xu, Zhou, and Wang (2000) write that at least 15 teeth are present, arranged like those in troodontids. There are no anterior serrations on the teeth. However, there are serrations on the posterior teeth. Basal constriction is found on the posterior teeth, and the crown is less compressed. The constriction is between the root and the crown. The authors mention those two features as bird-like and write that M. zhaoianus presents, “the more bird-like teeth” (Xu, Zhou, and Wang 2000).
No teeth are mentioned for M. gui. For M. hanqingi, the number of teeth cannot be certain. However, the authors write that it may be 16. Four teeth are preserved in position. There are minute serrations in one tooth (Gong et al. 2012). The authors of the original publication of M. hanqingi (Gong et al. 2012) mention that the serration pattern varies in the skulls known of Microraptor. They also write that serration is ambiguous in their specimen (Gong et al. 2012).
The presence of teeth is not problematic for Microraptor. Teeth have been noted as a dinosaurian feature, as modern birds do not possess teeth. However, some extinct birds have teeth like Hesperornis and Ichthyornis (Martin and Stewart 1977), Yanornis and Yixianornis (Zhou and Zhang 2001), and Sapeornis (Wang et al. 2017).
Regarding serration, as it is now known, no existing literature mentions serrated teeth in extinct birds. Also, heterodonty (different types of teeth) is not yet known for extinct birds.
Postorbital
No postorbital information is mentioned for M. zhaoianus. A triradiate postorbital was identified in M. gui and M. hanqingi. For M. gui, “Little can be said about the cranial morphology but a tri-radiate postorbital is identifiable” (Xu et al. 2003). And for M. hanqingi, “The holotype of M. gui has a skull, but the anterior portions are dubious and the posterior cranial elements are too badly crushed to reconstruct the cranium although it confirms the presence of a tri-radiate postorbital in this genus. The postorbital of M. hanqingi is similar” (Gong et al. 2012).
This feature is not found in modern birds (Bhullar et al. 2016) but is present in many dinosaurs (Nesbitt 2011). The presence of a postorbital was also confirmed in a Microraptor specimen found in China in 2014 (Pei et al. 2014). However, although extant birds do not possess a postorbital, they have it as an embryo. This bone fuses with the frontal before hatching. This fusion occurs during embryonic development in birds (Smith-Paredes et al. 2018). Thus, the presence of an unfused postorbital may not be a diagnostic feature for dinosaurs. In fact, recent research shows that fossil forms of several taxa of birds and dinosaurs matured much more slowly, having had longer gestation times than their modern counterparts. For example, Yang and Sander (2018) argued that the reason modern birds no longer develop teeth is because they spend less time in development than their fossil forebears. Though, as yet speculative, longer development and slower maturation times might have led to a longer time for the postorbital to fuse.
That is, it is possible that the slower development observed in some fossil taxa allowed the postorbital bone to remain unfused for a longer duration, potentially persisting into post-hatching stages. Raising the possibility that an unfused postorbital may reflect differences in developmental timing rather than a fundamental taxonomic distinction.
Smith-Paredes et al. (2018; Supplementary Figure 9) also state regarding the postorbital:
A postorbital has been found in a few enantiornithes [sic] preserving the temporal region. In many others, the area is crushed beyond recognition, which prevents identification of the element, but was probably present as inferred by phylogenetic bracketing. However, in some members of Enantiornithes, the postorbital is reduced (see Rapaxavis in O’Connor and Chiappe 2011), which suggests some taxa could have lost it in parallel to Euornithes. In the lineage leading to modern birds, an independent postorbital was lost at some point between the origin of Euornithes and the origin of Ornithurae; this cannot be established more precisely because preservation of basal Euornithes does not allow recognition of the presence or absence of a postorbital.
Although an evolutionary worldview is being applied to interpret the data, the citation above demonstrates that a postorbital is found in extinct birds. Enantiornithes are extinct birds with teeth and various features, including a postorbital. Modern birds do not exhibit many of the characteristics of Enantiornithes. The postorbital seems to be lost in the group (Euornithes) that includes the modern birds (Neornithes). Supporting these observations, Wellnhofer (2009) mentions the presence of a postorbital in an enantiornithine bird from the Lower Cretaceous in Spain. He also mentioned Confuciusornis from the Lower Cretaceous of China and wrote that this bird had a distinct postorbital.
O’Connor and Chiappe (2011) state that, when discussing the postorbital bone, “This element—absent among modern birds—is retained within Enantiornithes but is only definitively preserved in three specimens: LP 4450, Pengornis and Shenqiornis. In LP 4450 and Shenqiornis, it exhibits a typical T-shaped design, but in Pengornis, the caudodorsal (squamosal) ramus is either absent or broken. The postorbital of these specimens shows considerable variation in size and morphology.” A postorbital is also present in Navaornis hestiae, found in the Cretaceous of Brazil. This new genus and species was identified as Enantiornithes (Chiappe et al. 2024).
Additional evidence of a postorbital is present in extinct birds (fig. 1). A complete, well-preserved postorbital in Jeholornis prima, a bird from the Early Cretaceous of Jehol, China, exhibits a triradiate configuration (Hu et al. 2022). Also, Sapeornis is a bird from the Early Cretaceous in China. According to its description, it features a Y-shaped postorbital (Zhou and Zhang 2003). Furthermore, Yuanchuavis, a bird from the Early Cretaceous of China, also presents a triradiate postorbital (Wang et al. 2022).
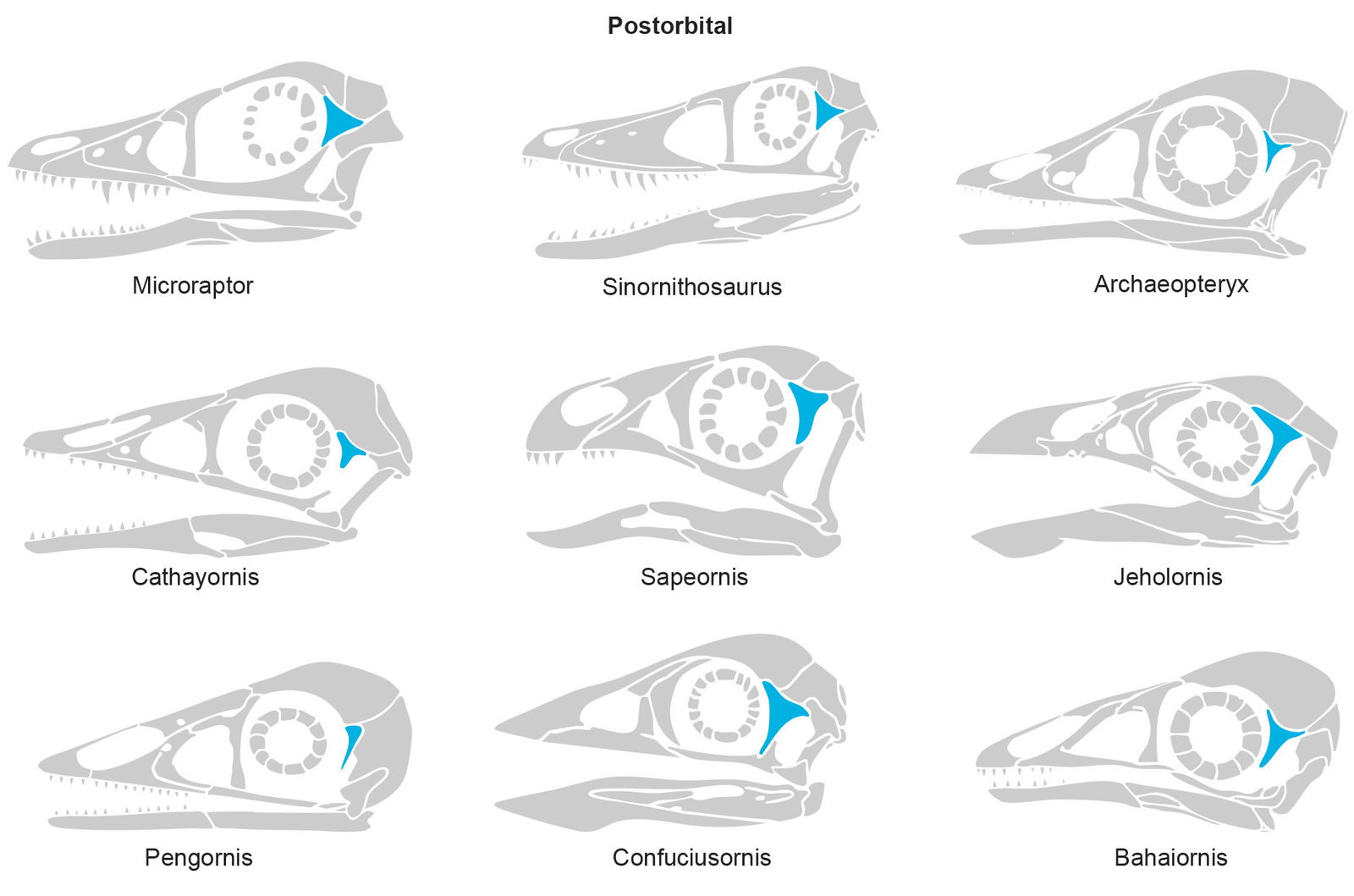
Fig. 1. Microraptor and Sinornithosaurus adapted from Hartman (n.d.) and Paul (2016) reconstructions. Hesperornis adapted from Everhart (2011). Archaeopteryx skull adapted from Wellnhofer (2009). Cathayornis, Confuciusornis, and Sapeornis adapted from Hartman (n.d.) reconstructions and Wang et al. (2017). Jeholornis adapted from Hu et. al. (2023). Pengornis and Bahaiornis adapted from Zelenkov (2017). Artwork by Joel Leineweber, used by permission.
It is also important to note that the presence or absence of a postorbital process might not indicate reptilian status for the reasons listed above. Postorbital bones are not always present in reptiles and thus should not be taken as a reptilian feature for Microraptor. The Amphisbaenia tribe, which includes burrowing worm-like or snake-like lizards, does not present a postorbital bone (Williston and Gregory 1925). It is also excluded from the extinct reptiles Araeoscelis and Hyperodapedon (Williston and Gregory 1925). Additionally, the postorbital bar is sometimes incomplete in lizards, snakes, and therapsids (Williston and Gregory 1925).
Moreover, Rauhut (2013) discusses the presence of a triradiate postorbital configuration in a new reconstruction of the Archaeopteryx skull. Although the article does not mention the word “triradiate,” the skull reconstruction in Fig. 5 of the publication reveals a triradiate structure for the postorbital. He uses this new reconstruction to support a more theropod-like skull for Archaeopteryx. The author also cites that a bird, Confuciusornis, has a postorbital structure. Even though he does not mention whether it is a triradiate structure. Hu et al. (2020) cite the well-developed postorbital in Confuciusornis. He also mentions that some Enantiornithes preserve some of the features found in Archaeopteryx. Archaeopteryx’s status as a bird has been disputed based less on anatomy and more on evolutionary bias, as Haynes (2022, 2023) has demonstrated.
The postorbital configuration is related to the kinesis (movement/mobility) of the skull of birds and dinosaurs (Wang and Hu 2017). Wang et al. (2022) cite Bock (1964) and Zusi (1984) and say that “avian cranial kinesis has been demonstrated to have improved feeding performance by increasing biting force, jaw closing speed, and food handling precision.”
Finally, Rahut (2013) notes that the possibility of the absence of a postorbital bar was, “one of the key arguments that led Bühler (1985) to propose a bird-like cranial kinesis in Archaeopteryx.” However, Hu et al. (2020) mention that “the skull of Sapeornis [an extinct bird] was largely akinetic, as in non-avian dinosaurs and palaeognaths (Hu et al. 2019).” Then, on the postorbital, it seems reasonable to conclude that the presence of a closed postorbital and an akinetic (immobile) skull can be features of a bird. Fig. 1 shows the list of extinct birds that present a postorbital bone.
Despite the evolutionary perspectives, extensive paleontological and anatomical data demonstrate that postorbital bones existed in multiple extinct bird and reptile taxa. Their presence or absence is subject to considerable developmental, functional, and biomechanical variability, cautioning against oversimplified interpretations of their taxonomic significance.
Lacrimal
The other feature of the cranial element is the lacrimal bone that is part of the orbit. This bone is found in one species (M. hanqingi). The feature of this bone for this species is described as, “The lacrimal is T-shaped and about 19.5 mm tall. The anterior ramus is 10.7 mm long; the posterior ramus is much shorter” (Gong et al. 2012).
However, extinct birds, such as those in the Enantiornithes group, exhibit the same T-shaped bone. Wellnhofer (2009) also mentions the lacrimal bone in Archaeopteryx, which has a T-bone shape. He explains that the mobility this bone configuration allows is in line with the prokinetic condition (independent motion of the upper beak relative to the rest of the skull) of the skull, as described by Bühler (1985). That is a condition present in most modern birds. However, Elzanowski (2002) mentioned that Wellnhofer (2009) suggested rhynchokinesis (ability to flex the upper part of the beak independently from the rest of the skull), but due to the Archaeopteryx’s upper jaw, its mobility might differ from the modern cranial kinesis categories. A T-shaped bone is also identified by Chiappe et al. (2024) as a lacrimal bone in Enantiornithes, specifically Navaornis hestiae.
Clarke (2009), analyzing two birds (Apatornis and Ichthyornis), discusses the presence of lacrimal and states, “In Aves, the lacrimal can articulate with (or coossify to) both the nasal and frontal, but may articulate exclusively with either of these two bones (Cracraft 1968). In Struthio camelus and Rhea americana, for example, the lacrimal usually contacts only the nasal (Cracraft 1968), unlike the condition in Ichthyornis dispar, while in tinamous, Apteryx, and Gallus gallus, it contacts the frontal and nasal (Cracraft 1968). In Anhimidae, it also articulates exclusively with the nasal (Cracraft 1968). Hesperornis regalis has the lacrimal primarily contacting the nasal, although also contacting the anterolateral portion of the frontal (Bühler, Martin, and Witmer 1988).” As shown above, a T-shape is necessary to articulate or touch the two bones (nasal and frontal), rather than just articulating with the jugal in a bar shape.
O’Connor and Chiappe (2011) state on a T-shaped lacrimal in two birds: “The morphology of this bone is definitively clear in only two specimens: Pengornis (IVPP V15336) and LP 4450.” Yuanchuavis also presents a T-shaped lacrimal, as shown in Wang et al. (2022). Sapeornis is also a bird that presents a T-shaped lacrimal bone, as described by Hu et al. (2020). The authors also mention that the Sapeornis’ lacrimal is like, “Archaeopteryx and some enantiornithines like Pengornis, Parapengorn and Pterygornis in overall shape.” The same happens to Cratonavis, a bird from the Early Cretaceous of China that presents a T-shaped lacrimal (Li et al. 2023).
A new species of fossil bird, belonging to the Anseriformes, was discovered in the Late Cretaceous system of Antarctica. Vegavis iaai was identified and described based on a nearly complete skull. This species also presents a T-shaped lacrimal. Little is known about this bone, except that it remains unfused. However, the figure on page 148 of the article shows a lacrimal with a T shape. (Torres et al. 2025). As shown in fig. 2, several extinct birds share the same features of the lacrimal found in the Microraptor. Fig. 2 shows the extinct birds that present a lacrimal bone.
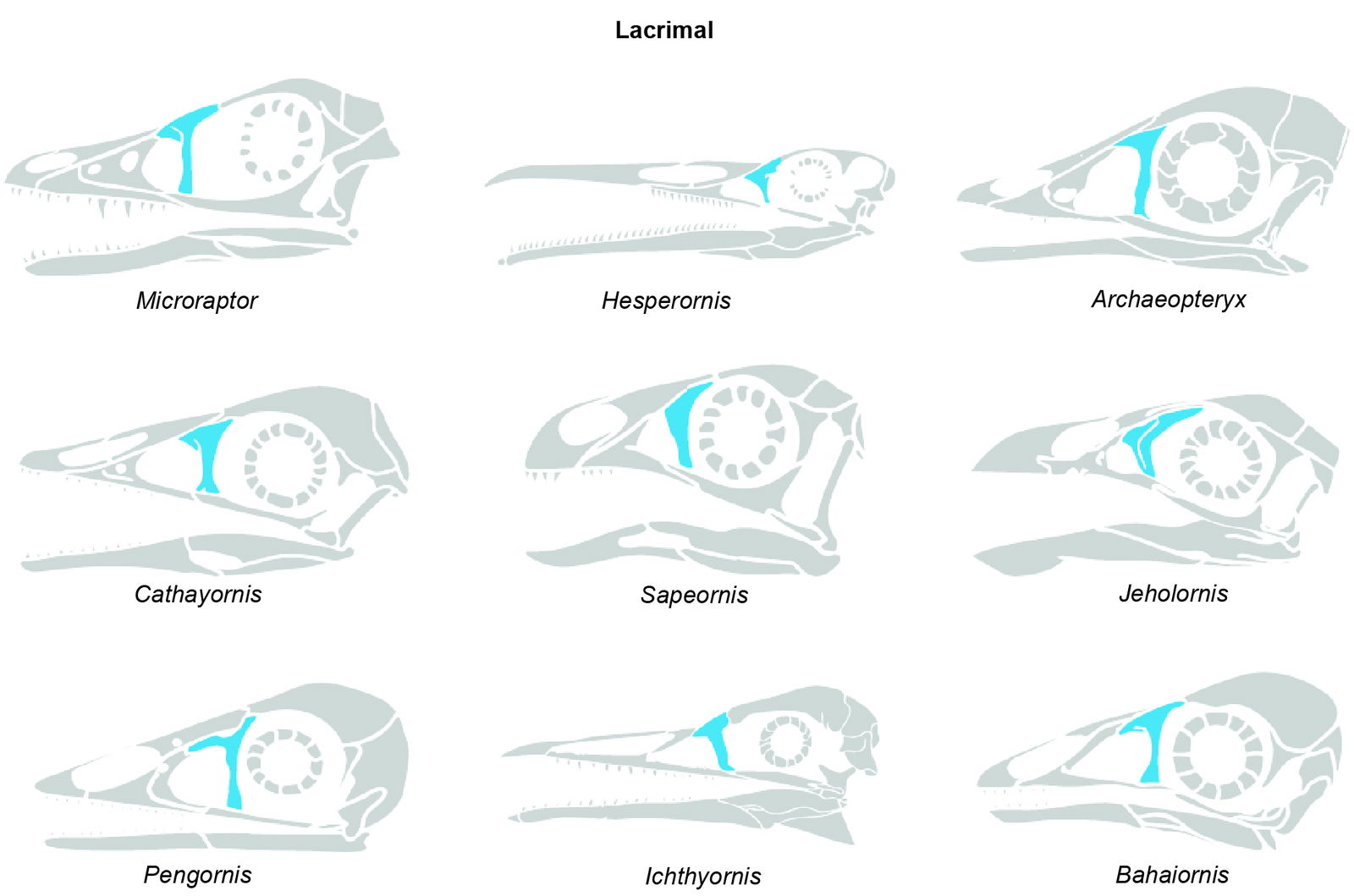
Fig. 2. Microraptor and Sinornithosaurus adapted from Hartman (n.d.) and Paul (2016) reconstructions. Hesperornis adapted from Everhart (2011). Archaeopteryx skull adapted from Wellnhofer (2009). Cathayornis, Confuciusornis, and Sapeornis adapted from Hartman (n.d.) reconstructions and Wang et al. (2017). Jeholornis adapted from Hu et. al. (2023). Pengornis and Bahaiornis adapted from Zelenkov (2017). Artwork by Joel Leineweber, used by permission.
Nasals
The characteristics of the nasal bones are cited in one species, M. hanqingi, “The nasals are 35 mm long and are thin, paired elements that widen anteriorly and posteriorly. They are widest (9.2 mm) across the anterior portion of the bone just before the concave border of the nares” (Gong et al. 2012). This aligns with a lightweight cranial structure. In contrast, in dinosaurs, nasal bones were often involved in supporting strong jaw muscles or forming crests for display or combat. The characteristics cited for the nasal bones of M. hangingi point to a bird-like feature. The nasal bones do not conform to a dinosaurian structure for this genus.
Quadrate
A quadrate (a bone that helps to connect the lower jaw to the rest of the skull) is mentioned for M. zhaoianus (Xu, Zhou, and Wang 2000), but it is not described in any of the specimens of those three species (M. zhaioanus, M. gui, and M. hanqingi). Wellnhofer (2009) mentions the single-headed feature as theropod-like and the double-headed feature as bird-like. However, a typical enantiornithine skull presents a single-headed quadrate (O’Connor and Chiappe 2011). Showing that a quadrate with that feature should not point to a reptilian-like skull.
It is also important to note that the word “theropod” has changed. When its definition was created, it only referred to dinosaurs. After 1985/1986, due to evolutionary influences, the term “theropod” began to include both dinosaurs and birds (Haynes 2022, 2023).
To summarize the anatomical information discussed in the previous sections and present a more straightforward comparison among the three described species of Microraptor (M. zhaoianus, M. gui, and M. hanqingi). Table 1 shows structured information of their cranial features. This comparative overview outlines the elements mentioned or described in each of the first three original publications of the species and contextualizes the significance of their preserved bones based on current paleontological literature. The table highlights patterns that may challenge the prevailing interpretations regarding Microraptor.
| Feature | M. zhaoianus | M. gui | M. hanqingi | Notes / Interpretation |
|---|---|---|---|---|
| Premaxilla | Present; sloping anterior margin (like Archaeopteryx, Sinornithosaurus) | Not mentioned | Remnants preserved | Sloping premaxilla considered bird-like |
| Maxilla | Contributes to external naris; similarity to Archaeopteryx and troodontids | Not mentioned | Present; 43.5mm; 14 alveoli; 4 teeth preserved | Longer and more recurved teeth in the maxilla than the dentary |
| Dentary | At least 15 teeth; packed like troodontids | Not mentioned | ~16 alveoli; teeth short, broad, uniformly sized | Tooth shape and packing varies; posterior serrations ambiguous |
| Tooth Serration | No anterior serrations; posterior serrations present; basal constriction | Not mentioned | One tooth with minute serrations; pattern ambiguous | Tooth serration has not been described in extinct birds |
| Postorbital | Not mentioned | Triradiate postorbital | Triradiate postorbital | Present in extinct birds like Jeholornis, Confuciusornis |
| Lacrimal | Not mentioned | Not mentioned | T-shaped; 19.5mm tall | T-shapes also found in Archaeopteryx, Sapeornis, enantiornithines |
| Nasal Bones | Mentioned but not described | Not mentioned | Thin, paired, widen anteriorly and posteriorly | Gracile form aligns with avian-like cranial lightness |
| Quadrate | Mentioned but not described | Not mentioned | Not mentioned | Avian/dinosaurian classification is inconclusive without it |
Table 1. Cranial morphological features analyzed of the three described species of Microraptor.
Conclusion
The anatomical analysis of Microraptor presented in this study questions the prevailing view that it was a feathered dinosaur and instead supports its classification as an extinct bird. The assumption that Microraptor must be a dinosaur based on certain skull features is weakened by facts showing that these traits also exist in extinct birds and even in some modern bird embryos. All the features examined on the skull of the first three original publications of Microraptor species show similarities with those of known extinct birds. The presence of multiple traits in Microraptor that are also found in birds suggests that Microraptor is a bird, not a dinosaur with feathers. Although the skull features might not be considered conclusive (since the features analyzed might not be diagnostic) for the status of Microraptor as a bird, the results in this paper seem to weigh in favor of it. This conclusion is particularly relevant considering the ongoing debate about the nature of “feathered dinosaurs.” Understanding Microraptor as an extinct bird, rather than a dinosaur with feathers, may help us recognize many other supposed feathered dinosaurs as just extinct birds.
Future research is planned to include postcranial characteristics. This study contributes to a growing body of information suggesting that our understanding of Mesozoic birds might be blurred with evolutionary influence. Moreover, many of the so-called feathered dinosaurs are, in fact, birds.
References
Alexander, David E., En-Pu Gong, Larry D. Martin, David A. Burnham, and Amanda R. Falk. 2010. “Model Tests of Gliding with Different Hindwing Configurations in the Four-Winged Dromaeosaurid Microraptor gui.” Proceedings of the National Academy of Sciences USA 107, no. 7 (January 25): 2972–2976.
Bechly, Günter. 2024a. “Fossil Friday: New Study Confirms ‘Feathered Dinosaurs’ were Secondarily Flightless Birds.” April 5. https://evolutionnews.org/2024/04/fossil-friday-new-study-confirms-feathered-dinosaurs-were-secondarily-flightless-birds/.
Bechly, Günter. 2024b. “Fossil Friday: More Evidence That ‘Feathered Dinosaurs’ were Secondarily Flightless Birds.” September 13. https://evolutionnews.org/2024/09/fossil-friday-more-evidence-that-feathered-dinosaurs-were-secondarily-flightless-birds/.
Bhullar, Bhart-Anjan S., Jesús Marugán-Lobón, Fernando Racimo, Gabe S. Bever, Timothy B. Rowe, Mark A. Norell, and Arhat Abzhanov. 2012. “Birds have Paedomorphic Dinosaur Skulls.” Nature 487, no. 7406 (May 25): 223–226.
Bhullar, Bhart-Anjan S., Michael Hanson, Matteo Fabbri, Adam Pritchard, Gabe S. Bever, and Eva Hoffman. 2016. “How to Make a Bird Skull: Major Transitions in the Evolution of the Avian Cranium, Paedomorphosis, and the Beak as a Surrogate Hand.” Integrative and Comparative Biology 56, no. 3 (September): 389–403.
Bock, Walter J. 1964. “Kinetics of the Avian Skull.” Journal of Morphology 114, no. 1 (January 1): 1–41.
Bühler, P. 1985. “On the Morphology of the Skull of Archaeopteryx.” In The Beginnings of Birds. Edited by M. K. Hecht, J. H. Ostrom, G. Biohl, and P. Wellnhofer, 135–140. Eichstätt, Germany: Freunde des Jura-Museums.
Bühler, Paul, Larry D. Martin, and Lawrence M. Witmer. 1988. “Cranial Kinesis in the Late Cretaceous Birds Hesperornis and Parahesperornis.” Ornithology 105, no. 1 (January 1): 111–122.
Chiappe, Luis M., Guillermo Navalón, Agustín G. Martinelli, Ismar De Souza Carvalho, Rodrigo Miloni Santucci, Yun-Hsin Wu, and Daniel J. Field. 2024. “Cretaceous Bird from Brazil Informs the Evolution of the Avian Skull and Brain.” Nature 635, no. 8038 (October 30): 376–381.
Clarke, Julia A. 2004. “Morphology, Phylogenetic Taxonomy, and Systematics of Ichthyornis and Apatornis (Avialae: Ornithurae).” Bulletin of the American Museum of Natural History 286 (August 27): 1–179.
Cracraft, Joel. 1968. “The Lacrimal-Ectethmoid Bone Complex in Birds: A Single Character Analysis.” The American Midland Naturalist 80, no. 2 (October 1): 316–359.
Elzanowski, Andrzej. 2002. “Archaeopterygidae (Upper Jurassic of Germany).” In Mesozoic Birds: Above the Heads of Dinosaurs. Edited by Luis M. Chiappe and Lawrence M. Witmer, 129–159. Berkeley, California: University of California Press.
Everhart, Michael J. 2011. “Rediscovery of the Hesperornis regalis Marsh 1871 Holotype Locality Indicates an Earlier Stratigraphic Occurrence.” Transactions of the Kansas Academy of Science 114, nos. 1–2 (April 1): 59–68
Feduccia, Alan. 2020. Romancing the Birds and Dinosaurs: Forays in Postmodern Paleontology. Irvine, California: Brown Walker Press.
Felice, Ryan N., Akinobu Watanabe, Andrew R. Cuff, Michael Hanson, Bhart-Anjan S. Bhullar, Emily R. Rayfield, Lawrence M. Witmer, Mark A. Norell, and Anjali Goswami. 2020. “Decelerated Dinosaur Skull Evolution with the Origin of Birds.” PLoS Biology 18, no. 8 (August 18): e3000801.
Field, Daniel J., Michael Hanson, David Burnham, Laura E. Wilson, Kristopher Super, Dana Ehret, Jun A. Ebersole, and Bhart-Anjan S. Bhullar. 2018. “Complete Ichthyornis Skull Illuminates Mosaic Assembly of the Avian Head.” Nature 557, no. 7703 (May 2): 96–100.
Gong, En-Pu, Larry D. Martin, David A. Burnham, Amanda R. Falk, and Lian-Hai Hou. 2012. “A New Species of Microraptor from the Jehol Biota of Northeastern China.” Palaeoworld 21, no. 2 (May 22): 81–91.
Hartman, Scott. n.d.. “Theropod Skeletal Reconstructions.” Dr. Scott Hartman’s Skeletal Drawing.com. https://www.skeletaldrawing.com/theropods/.
Haynes, Gabriela. 2022. “The Debate over Classification of Archaeopteryx as a Bird.” Answers Research Journal 15 (September 14): 285–300. https://answersresearchjournal.org/dinosaurs/debate-classification-archaeopteryx-bird/.
Haynes, Gabriela. 2023. “Reply to the ‘Response to “The Debate over Classification of Archaeopteryx as a Bird”’.” Answers Research Journal 16 (December 6): 595–634. https://answersresearchjournal.org/dinosaurs/reply-response-debate-classification-archaeopteryx-bird/.
Hu, Han, Jingmai K. O’Connor, Paul G. McDonald, and Stephen Wroe. 2020. “Cranial Osteology of the Early Cretaceous Sapeornis chaoyangensis (Aves: Pygostylia).” Cretaceous Research 113 (May 12): 104496.
Hu, Han, Yan Wang, Matteo Fabbri, Jingmai K O’Connor, Paul G. McDonald, Stephen Wroe, Xuwei Yin, Xiaoting Zheng, Zhonghe Zhou, and Roger B. J. Benson. 2023. “Cranial Osteology and Palaeobiology of the Early Cretaceous Bird Jeholornis prima (Aves: Jeholornithiformes).” Zoological Journal of the Linnean Society 198, no. 1 (October 24): 93–112.
Hu, Han, Gabriele Sansalone, Stephen Wroe, Paul G. McDonald, Jingmai K. O’Connor, Zhiheng Li, Xing Xu, and Zhonghe Zhou. 2019. “Evolution of the Vomer and Its Implications for Cranial Kinesis in Paraves.” Proceedings of the National Academy of Sciences USA 116, no. 39 (September 9): 19571–19578.
Li, Zhiheng, Min Wang, Thomas A. Stidham, and Zhonghe Zhou. 2023. “Decoupling the Skull and Skeleton in a Cretaceous Bird With Unique Appendicular Morphologies.” Nature Ecology and Evolution 7, no. 1 (January 2): 20–31.
Martin, Larry D., and J. D. Stewart. 1977. “Teeth in Ichthyornis (Class: Aves).” Science 195, no. 4284 (March 25): 1331–1332.
Nesbitt, Sterling J. 2011. “The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades.” Bulletin of the American Museum of Natural History 2011, no. 352 (April 29): 1–292.
O’Connor, Jingmai K., and Luis M. Chiappe. 2011. “A Revision of Enantiornithine (Aves: Ornithothoraces) Skull Morphology.” Journal of Systematic Palaeontology 9, no. 1 (February 28): 135–157.
Paul, Gregory S. 2016. The Princeton Field Guide to Dinosaurs. 2nd ed. Princeton, New Jersey: Princeton University Press.
Pei, Rui, Quanguo Li, Qingjin Meng, Ke-Qin Gao, and Mark A. Norell. 2014. “A New Specimen of Microraptor (Theropoda: Dromaeosauridae) From the Lower Cretaceous of Western Liaoning, China.” American Museum Novitates 2014, no. 3821 (December 22): 1–28.
Rauhut, Oliver W. M. 2013. “New Observations on the Skull of Archaeopteryx.” Paläontologische Zeitschrift 88, no. 2 (June 7): 211–221.
Senter, Phil, Rinchen Barsbold, Brooks B. Britt, and David A. Burnham. 2004. “Systematics and Evolution of Dromaeosauridae (Dinosauria: Theropoda).” Bulletin of Gunma Museum of Natural History 8 (January): 1–20.
Smith-Paredes, Daniel, Daniel Núñez-León, Sergio Soto-Acuña, Jingmai O’Connor, João Francisco Botelho, and Alexander O. Vargas. 2018. “Dinosaur Ossification Centres in Embryonic Birds Uncover Developmental Evolution of the Skull.” Nature Ecology and Evolution 2, no. 12 (November 6): 1966–1973.
Torres, Christopher R., Julia A. Clarke, Joseph R. Groenke, Matthew C. Lamanna, Ross D. E. MacPhee, Grace M. Musser, Eric M. Roberts, and Patrick M. O’Connor. 2025. “Cretaceous Antarctic Bird Skull Elucidates Early Avian Ecological Diversity.” Nature 638, no. 8049 (February 5): 146–151.
Turner, Alan H., Peter J. Makovicky, and Mark A. Norell. 2012. “A Review of Dromaeosaurid Systematics and Paravian Phylogeny.” Bulletin of the American Museum of Natural History 2012, no. 371 (August 17): 1–206.
Wang, Min, and Han Hu. 2017. “A Comparative Morphological Study of the Jugal and Quadratojugal in Early Birds and Their Dinosaurian Relatives.” The Anatomical Record 300, no. 1 (December 21): 62–75.
Wang, Min, Thomas A. Stidham, Jingmai K. O’Connor, and Zhonghe Zhou. 2022. “Insight Into the Evolutionary Assemblage of Cranial Kinesis From a Cretaceous Bird.” eLife 11 (December 5). https://doi.org/10.7554/elife.81337.
Wang, Yan, Han Hu, Jingmai K. O’Connor, Min Wang, Xing Xu, Zhonghe Zhou, Xiaoli Wang, and Xiaoting Zheng. 2017. “A Previously Undescribed Specimen Reveals New Information on the Dentition of Sapeornis chaoyangensis.” Cretaceous Research 74 (June): 1–10.
Wellnhofer, Peter. 2009. Archaeopteryx: The Icon of Evolution. Munich, Germany: Verlag Dr. Friedrich Pfeil.
Williston, Samuel Wendell, and William King Gregory. 1925. The Osteology of the Reptiles. Cambridge, Massachusetts: Harvard University Press.
Wood, Tod, and Paul Garner. n.d. Let’s Talk Creation. https://www.youtube.com/watch?v=OgpNJpoJEOQ. minutes 57:43–58:20
Xu, Xing, Zhonghe Zhou, and Xiaolin Wang. 2000. “The Smallest Known Non-Avian Theropod Dinosaur.” Nature 408, no. 6813 (December 7): 705–708.
Xu, Xing, and Xiao-Chun Wu. 2001. “Cranial Morphology of Sinornithosaurus millenii Xu et al. 1999 (Dinosauria: Theropoda: Dromaeosauridae) From the Yixian Formation of Liaoning, China.” Canadian Journal of Earth Sciences 38, no. 12 (December 1): 1739–1752.
Xu, Xing, Zhonghe Zhou, Xiaolin Wang, Xuewen Kuang, Fucheng Zhang, and Xiangke Du. 2003. “Four-Winged Dinosaurs From China.” Nature 421, no. 6921 (January 28): 335–340.
Yang, Tzu-Ruei, and P. Martin Sander. 2018. “The Origin of the Bird’s Beak: New Insights From Dinosaur Incubation Periods.” Biology Letters 14, no. 5 (May 1): Article ID: 20180090.
Zelenkov, N. V. 2017. “Early Cretaceous Enantiornithine Birds (Aves, Ornithothoraces) and Establishment of the Ornithuromorpha Morphological Type.” Paleontological Journal 51, no. 6 (November 1): 628–642.
Zhou, Zhonghe, and Fucheng Zhang. 2001. “Two New Ornithurine Birds From the Early Cretaceous of Western Liaoning, China.” Chinese Science Bulletin 46, no. 15 (August 1): 1258–1264.
Zhou, Zhonghe, and Fucheng Zhang. 2003. “Anatomy of the Primitive Bird Sapeornis chaoyangensis From the Early Cretaceous of Liaoning, China.” Canadian Journal of Earth Sciences 40, no. 5 (May 1): 731–747.
Zusi, Richard L. 1984. “A Functional and Evolutionary Analysis of Rhynchokinesis in Birds.” Smithsonian Contributions to Zoology 395 (January 1): 1–40.








