
The views expressed in this paper are those of the writer(s) and are not necessarily those of the ARJ Editor or Answers in Genesis.
Abstract
The lycopod stumps at Fossil Grove in Victoria Park, Glasgow, are widely believed to be in situ in a sandy mudstone. However, flattened axial systems, unattached stigmarian appendages, and the thinness, lack of bioturbation and lack of soil development in the sandy mudstone suggest the stumps were emplaced allochthonously. The limited metamorphism and lack of bioturbation in Fossil Grove sediments not only confirm allochthony, but suggest all the sediments were deposited in the same diluvial catastrophe. The distribution of stumps with respect to possible megaripples in the underlying unit, combined with the deformation of axial systems, suggest the lycopod trees lived and grew in water, and decomposed while floating in water, before being deposited at this location. Subsurface borehole and the plunging synclinal outcrop pattern of sediments in the Glasgow region suggest Fossil Grove sediments sit atop thousands of feet of fossiliferous sediments and thousands of feet of flood basalts. Unlike Clarey and Tomkins’ (2016) claim, Fossil Grove stumps were allochthonously deposited well into the Flood.
When Clarey and Tomkins’ (2016) autochthony criteria are corrected, generalized, and expanded, 12 autochthony criteria applied to Fossil Grove stumps argue strongly for their allochthonous emplacement. Among Carboniferous fossil forests in general, their rapid and prolonged burial, the placement of most of them over coal, the commonness of a single higher taxon, and the near-ubiquity of internal-mold preservation collectively suggest that the application of the same 12 criteria to claims of in situ Paleozoic forests will indicate all of them were generated allochthonously during the Flood.
Introduction
Some creationists (such as Scheven 1981) have argued that pre-Flood arborescent lycopods grew floating on water. Wise (2003) even argued that arborescent lycopods were part of a biome that floated atop the pre-Flood ocean. Clarey and Tomkins (2016), in an effort to refute all claims of antediluvian floating forests, claim that the lycopod fossils at Fossil Grove in Victoria Park in Glasgow, Scotland are in situ stumps1 from a land-based lycopod forest. They argue that the Fossil Grove stumps were rooted in the same sandy mud now observed surrounding the axial systems (“roots”) of the fossil stumps as a sandy mudstone. Clarey and Tomkins (2016) further argue that this sandy mudstone lies atop pre-Flood bedrock on a structural high dating from pre-Flood times. This structural high was, they argue, a hill that was not covered with Flood sediment until after the deposition of all lower Paleozoic (Vendian through Lower Carboniferous) strata.
A visit by the author to the Fossil Grove locality, however, suggested that previous interpretations of the Fossil Grove site were in error, including the interpretation of Clarey and Tompkins (2016). This paper reviews the problems with autochthonous interpretations of the Fossil Grove fossils, and offers an alternative interpretation of Fossil Grove and other Paleozoic “fossil forests.”
Fossil Grove Strata
In 1885, during a depression in the Scottish shipbuilding industry, the commissioners of the burghs of Whiteinch and Partick leased 46 acres of land and provided jobs to unemployed shipbuilding laborers by having them construct a public park (Allison and Webster 2017; Gunning 1995). Work on the park began in November of 1886 (Gunning 1995). The park was officially opened on 2 July 1887, and was named Victoria Park at the approval of, and in honor of, Queen Victoria during her Golden Jubilee (Gunning 1995). Landscaping an abandoned whinstone (dolerite) quarry in the southeastern corner of the park continued after the park’s official opening. In the closing months of 1887, in a portion of the quarry where the main dolerite body had been completely quarried away, workers encountered a fossil stump (Young and Glen 1888). Recognizing the attraction that such fossils might have in a public park, excavation continued. First, four more stumps were excavated, and by late Spring, 1888, a total of 11 stumps had been revealed2 (Young and Glen 1888). Again recognizing the value such fossils might have in a public park, town commissioners hired the noted architect Peter MacGregor Chalmers (1859–1922) to design and build a building for the fossils (Gunning 1995; Glasgow City Council n.d.). After initially protecting the fossils with a temporary structure, the current brick-walled building was built in 1889, and opened 1 January 1890 (Allison and Webster 2017; Gunning 1995).
Description of Fossil Grove strata
Since the discovery of the fossils, Glasgow has grown around Victoria Park and the burghs of Whiteinch and Partick. A search of the area around the Fossil Grove site by the author revealed no current bedrock exposures for more than a mile around the locality in all directions. Thus direct observational research on Fossil Grove rocks is restricted to those exposed in and immediately around the Fossil Grove building.
Three lithologies are found at Fossil Grove—one igneous and two sedimentary. The igneous rocks of Fossil Grove are an alkali olivine dolerite—i.e., teschenite (Allison and Webster 2017; Hall, Browne, and Forsyth 1998). Roughly 17 ft of dolerite is exposed in the old quarry walls to the northwest of the Fossil Grove building. Thinner dolerite layers are exposed among the sedimentary rocks in the building—one of those layers cutting through Stumps 8 and 93(represented diagrammatically in fig. 1). The dolerite has chilled margins where it is in contact with sediments (Allison and Webster 2017; McCallien 1938, 162; Young and Glen 1888) and thin dolerite layers have compositions consistent with chemical alteration by carbon dioxide released from heated shales (Keen 1992; McCallien 1938, 162). The dolerite/sediment contacts parallel, and do not cross-cut, sedimentary layers.4 On geologic maps,5 laterally continuous bands of dolerite parallel formational contacts (see, e.g., the dolerite sills and nearby formational contacts in fig. 2). Young and Glen (1888) claimed the dolerites of the Glasgow region are found in all strata older than the Upper Red (Permian) sandstones—including all the sediments represented in fig. 3. The dolerites of the Glasgow region also consistently yield Lower Permian radiometric ages (Hall, Browne, and Forsyth 1998, 48).
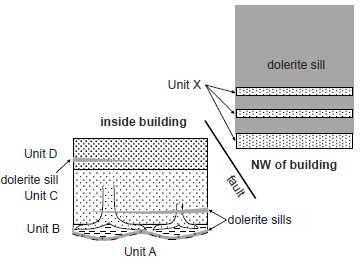
Fig. 1. Fossil Grove Strata. Representation of the strata of Fossil Grove to show stratigraphic relationships. Not to scale.
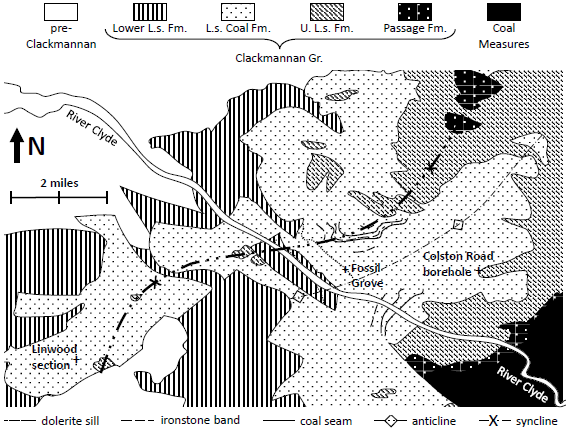
Fig. 2. Lower Limestone Formation of Glasgow. Bedrock geology from fig. 1 of Hall, Browne, and Forsyth 1998. Lower Limestone Formation sills, ironstone bands and coal seams from fig. FG2 of Allison and Webster 2017.
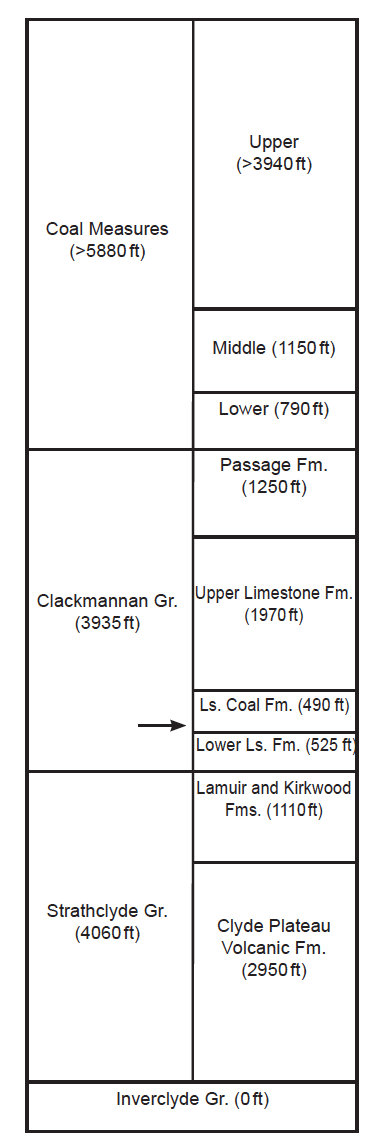
Fig. 3. Carboniferous Stratigraphy of Glasgow. Stratigraphic nomenclature of Hall, Browne, and Forsyth 1998. Formational thicknesses are from table 4 of Monaghan 2014, supplemented with this paper’s conclusion about the thicknesses of the Lower Limestone and Limestone Coal Formations. The arrow indicates the approximate stratigraphic location of Fossil Grove.
Based on his own examination of rocks of the site, this paper’s author divides the sedimentary rocks (silty shales and sandstones) at Fossil Grove into five stratigraphic units (Units A, B, C, D, and X), with the rock of Unit A not directly observed, but rather inferred from characteristics of what is most likely its upper boundary.
All but one of the stratigraphic units at Fossil Grove (Units D, C, B, and A) are found inside the Fossil Grove building (represented diagrammatically on the left side of fig. 1). Beginning with the uppermost of the units in the building, Unit D6 is a coarsening upward, climbing-rippled, cross-bedded, coarse siltstone with millimeter-scale, occasionally disordered, laminae (Gastaldo 1986). The only fossils reported from Unit C are disordered, coalified, compression megaflora, and they become less common up the unit (Gastaldo 1986).
What is here called Unit C lies conformably under Unit D (represented diagrammatically on the left side of fig. 1). Unit C is 35 in7 (Keen 1992) of fine- to medium-grained (Gastaldo 1986), grey (MacGregor and Walton 1948; McLean 1973) sandstone8 which grades upward from thick (>8 in), massive beds (Gastaldo 1986) with rare ripple-marked bedding surfaces (Gastaldo 1986; Keen 1992) to thinner beds (Keen 1992) with more ripple marks (Gastaldo 1986). Unit C ripple marks from the level of the top of the stumps (Gastaldo 1986) and those near Log C (R1 in fig. 4) are slightly asymmetrical, indicating a current flow to the southwest (Allison and Webster 2017; Gastaldo 1986). The most prominent fossils of Unit C are the Fossil Grove stumps. Even though the bases of the stumps are found in the underlying Unit B, each stump penetrates upward into the sandstone of Unit C. Each stump penetrates to a different height, and none of the stumps makes it to the top of Unit C (Gastaldo 1986; Keen 1992; Young and Glen 1888). All that currently remains of each Fossil Grove stump is actually sandstone of this unit (Gastaldo 1986; Young and Glen 1888). At the time of excavation, the sandstone of Unit C above each stump was continuous with the sandstone inside each stump and with the sandstone inside its axial structures. The sandstone of the stump and axial structures was originally surrounded by a thin layer of coalified material that sloughed off during the rigors of the excavation. Other fossils reported from Unit C include (1) unattached sand-filled stigmaria fragments (Gastaldo 1986; e.g., E in fig. 4), (2) logs or branches,9 (3) Cyperites compression fossils (Gastaldo 1986), and (4) Arenicola trace fossils (Young and Glen 1888). Most of the logs or branches are unidentified, but at least four of them are flattened Lepidodendron logs (A, B, C, and X in fig. 4) (Keen 1992; fig. FG6 of Allison and Webster 2017), with Logs A and C oriented in a NE–SW direction (Gastaldo 1986; see fig. 5 in this paper). The logs are at different levels in Unit C, most of them in the upper half of the unit, Log A being in the lower half and Log B being near the base of Unit C (or even, perhaps, in Unit B).
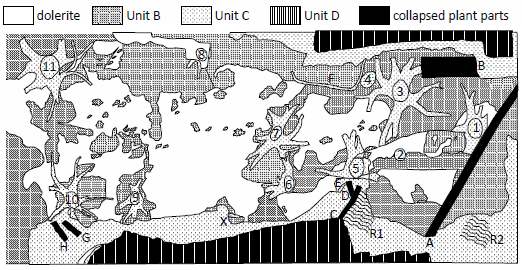
Fig. 4. Plan view of Fossil Grove. Lithotypes and most fossil locations from a map by Samuel Elder, Robert J. S. McCall, A. Kerr Pringle, and William J. Neaves published in fig. 79 of McCallien (1938). Stumps 2 and 4, the numbering, the lettering (through H) were from fig. 1 of MacGregor and Walton (1948), and the location of R1 was from fig. II of MacGregor and Walton (1948). Branch X location was from fig. 1 of MacGregor and Walton (1983) and R2 location was from D in fig. 2.1 of Keen (1992; not as indicated in fig. FG5 of Allison and Webster 2017). The area represented is 78 ft 8 in by 34 ft.
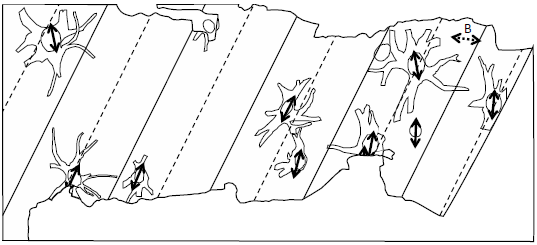
Fig. 5. Paleoflow Indicators. Unit A megaripples are indicated by solid lines (for crests) and dashed lines (for troughs). Dashed arrows indicate orientation of long axes of prostrate stems in Unit B. Solid arrows indicate orientation of long axes of stump cross sections, long prostrate stems and stigmaria, and current direction indicated by asymmetric ripples—all in Unit C. Orientations measured by author. The area represented is 78 ft 8 in by 34 ft.
What is here called Unit B is the lowest of the three units exposed in the Fossil Grove building and it conformably underlies Unit C (represented diagrammatically on the left side of fig. 1). Unit B is a carbonaceous, irregularly and thinly laminated, dark grey, silty mudstone10 which is currently exposed (1) along the base of the vertical walls where sediments were cut away to expose the stumps, (2) under a number of the stumps and axial systems, and (3) on a portion of the floor of the Fossil Grove building—especially along its eastern end (see fig. 4).11 The fossils reported from Unit B are “numerous fragments and impressions of plant remains” (Young and Glen 1888), “rare coalified compressions” (Gastaldo 1986, 78), Stigmaria ficoides (Kidston 1888, e.g., F in fig. 4), and stigmarian appendages (“rootlets”) cross cutting Unit B laminae—especially in the vicinity of Stump 11 (Gastaldo 1986; Keen 1992).
Unit B is currently sitting atop dolerite (represented diagrammatically on the left side of fig. 1). As Keen (1992) claims, this dolerite surface is undulatory. In fact, the author observed the undulations to be smooth, symmetric, and sinusoidal, with a wavelength of about 12 ft and an amplitude of about 1 ft. And, on the north wall, the lower part of the Unit B sandstone layer overlying this surface seems to mirror the undulatory pattern of the underlying dolerite, all the while showing no clear sediment thickness differences between the troughs and crests.12
The unit here called Unit X is the only sedimentary unit exposed outside the Fossil Grove building (represented diagrammatically on the right side of fig. 1). Exposures of Unit X are restricted to the walls of the deeper portion of the old quarry—all within about 100 ft of the northwest corner of the Fossil Grove building. Most of the walls of this part of the quarry are dolerite. Unit X is made up of several sandstone beds—most underlying the dolerite at the base of the quarry walls, but some higher up the quarry walls surrounded above and below by dolerite. The dip of the largest sandstone body of Unit X was measured to be at least 13° to the northeast. Although Unit X beds include up to 6 in of ½ in–2 in-thick laminar beds, most of the sandstones show medium- to thick-bedded (1.5 ft–3 ft) crossbedding. Allison and Webster (2017, 12–13) interpret one sandstone of Unit X as displaying a channel with a SW/NE orientation. However, it was this author’s impression that Allison and Webster’s “channel” does not cross cut the laminae of the underlying sandstone as Allison and Webster claim, and as would be expected with a channel. Rather, the sides of the channel appear to be bounding surfaces of steeply dipping sandstone laminae that gradually thicken as they rise upward. This suggests the channel might actually be formed by very steep cross-beds in a thick (>3 ft) sandstone bed. This interpretation would also suggest a current direction orthogonal to the current direction indicated by Allison and Webster’s channel. Though lying 10–15 ft below the plane of the Fossil Grove building floor, Unit X strata underlie at least 17 ft of dolerite—through which the plane of the Fossil Grove building floor projects.
Interpretation of Fossil Grove strata
The sedimentary rocks at Fossil Grove have always been placed together in the same named unit.13 In the most recent nomenclature (of 1996), they are part of the Limestone Coal Formation of the Clackmannan Group (Hall, Browne, and Forsyth 1998). They lie just beneath more than a mile of coal-bearing sediments of the Glasgow region (coals are found from the Limestone Coal Formation through the Coal Measures in fig. 3).
Regarding Fossil Grove dolerite, since it is an igneous rock, has both chilled margins and chemical alteration near contacts with sedimentary rocks, and parallels rather than cross cuts the sedimentary bedding, the Fossil Grove dolerites are interpreted as sills. The cross cutting of Fossil Grove stumps, the Lower Permian radiometric ages, and intrusion into sediments even thousands of stratigraphic feet higher, indicates that the Fossil Grove dolerite was intruded long after the deposition of the Fossil Grove sediments and their contained fossils.
Understanding the dolerite as a later sill intrusion very much affects the interpretation of Unit X. Although dolerite sills are claimed to intrude all pre-Permian rocks in the Glasgow region, thick sills are uncommon, often separated from other thick sills by hundreds of feet of sediment. At Fossil Grove, more than 15 ft of dolerite is exposed over a distance of nearly 2 mi (see fig. 2), and sills of similar thickness are hundreds of stratigraphic feet away. This suggests that the main dolerite body that was quarried away from above the fossil trees in the nineteenth century is the same thick dolerite sill currently exposed to the northwest of the Fossil Grove building. Since a projection of the dip of the floor of the Fossil Grove building would pass through the middle of the sill to the northwest, Allison and Webster (2017, 11) reason that a fault must lie between the Fossil Grove building and the quarry exposures to the northwest. The fault has at least 6 ft of vertical displacement, with the Fossil Grove building on the up-thrown side of the fault. With the fault not visible, the exact position of the quarried-away dolerite unknown, and an unknown amount of strike-slip, the exact displacement is not known. This means that the exact stratigraphic position of Unit X relative to the sediments in the Fossil Grove building is not known. Unit X looks very similar to Unit D, and could be at the same stratigraphic level or a few feet above, but the faulting renders the correlation uncertain. Furthermore, since Unit X beds are relatively thin beds under, or in the midst of, very much thicker sill rocks, they might not simply be a series of sedimentary beds vertically separated from one another by sills. They might, instead, be laterally extensive but discontinuous lithosomes suspended as xenoliths in a thick dolerite sill. The latter is suggested by much larger dip of the largest sedimentary body of Unit X compared to the dip of the sediments in the Fossil Grove building. Since lithosomes in lava could have moved in any direction (laterally, vertically, rotationally), even the relative position and orientation of individual Unit X sedimentary bodies to each other is unknown. Based on the crossbeds of Unit X, the most certain conclusions that can be drawn are that Unit X sands were deposited under conditions of substantial current. As with all the beds at Fossil Grove, Unit X sediments need a more careful examination, but they appear to be subaqueous dune or sand wave deposits stratigraphically at or above Unit D—the uppermost sediments exposed in the Fossil Grove building.
Understanding the dolerite as a sill is also important in inferring something about the sediment underlying Unit B. Most of the floor of Fossil Grove between the stumps, especially in the middle of the building, is actually dolerite (Allison and Webster 2017; Keen 1992; MacGregor and Walton 1948; McCallien 1938; see also fig. 4). Since Unit B sits atop the dolerite sill, and the dolerite does not seem to crosscut the basal laminae of Unit B, the most natural interpretation is that the dolerite surface preserves the basal bedding surface of Unit B before the intrusion of the dolerite. It may be possible that a sill could generate an undulatory surface by intruding magma with laterally varying thickness. However, the author has never seen a sill with such a cross-section, and it is not clear what sort of process could generate as smooth, regular, and symmetric pattern as is found on the floor of the Fossil Grove building. Furthermore, with the Fossil Grove sediments buried under more than a mile of sediment at the time of the intrusion, it is not clear how such differences in intrusion thickness would be permitted at all. It is most probable, then, that the dolerite surface is preserving the basal bedding surface of Unit B at the time of deposition of Unit B. The undulatory surface under Unit B would then also be the undulatory surface of the stratigraphic unit underlying B—the unobserved stratigraphic unit here called Unit A.14
The dolerite, then, is following the bedding surface of large, parallel ripple marks in Unit A. The orientation of these megaripples would indicate a substantial NW/SE current during the deposition of Unit A (see fig. 5). The symmetric nature of the megaripples combined with constant sediment thickness in basal Unit B across crests and troughs further suggest that upper Unit A and lower Unit B were likely emplaced with bidirectional currents. It is even possible that Units A and B are not separate units, but lower and upper layers from the same depositional event (only later separated by the dolerite intrusion). Bidirectional currents of the magnitude necessary to generate megaripples of that size may indicate tsunami or tsunami-like deposits.
Unit B is usually interpreted as the sediment in which the fossil trees were originally rooted (Allison and Webster 2017; Clarey and Tomkins 2016; Cleal and Thomas 1995; Gastaldo 1986; Gunning 1995; Keen 1992; Lawson and Lawson 1976, 71; MacFadyen and Gordon 2006, 14–15; MacGregor and Walton 1948; McLean 1973; Owen et al. 2007; Young and Glen 1888). Unit B is thus interpreted as a paleosol (Clarey and Tomkins 2016; Gastaldo 1986; Gunning 1995; Keen 1992; Owen et al. 2007). In the particular interpretation of Clarey and Tomkins (2016), Units C and D were rapidly formed and buried during the Flood, whereas Unit B was formed before the Flood and persisted for a considerable time period prior to the Flood. However, there are problems with a paleosol interpretation of Unit B. MacGregor and Walton (1948), for example, postulated that the original soil around the trees may have been washed away prior to the deposition of the sediment that encased and killed the trees. Although MacGregor and Walton did not give their rationale for this claim, it suggests that they saw no evidence of soil development in Unit B and/or the thickness of Unit B was insufficient for it to be the rooting soil for the original stumps. In fact, the first problem with Unit B as a paleosol is that no one has reported any evidence of soil development in it. Second, the thickness of Unit B seems inadequate. The author’s observation of Unit B for most of the Fossil Grove stumps15—and consistent with the cross-sections in fig. 80 of McCallien (1938)—was that it is only a few inches thick—at most originally barely covering the stigmaria two bifurcations from the trunk, let alone covering the stigmaria near the trunks.16 However, with rootlet scars on all sides of stigmaria, rootlets must have extended upward from the tops of stigmaria—at least inches and perhaps multiple feet (as fossil stigmaria can be that long). Thus the existing sediments of Unit B are by no means thick enough to have been the soil—or at least the entire depth of soil—in which the stumps were rooted. A third problem with the paleosol interpretation is that the stigmarian appendages that Gastaldo (1986) reported from this unit—even though they are claimed to cross cut the laminae—are not attached to stigmaria, let alone the axial systems of the known stumps. Thus, an allochthonous origin cannot be ruled out for them as well. Fourthly, the laminae of Unit B appear to be depositional laminae, unmodified by soil development. Fifthly, as noted by Gastaldo (1986), the axial systems of the Fossil Grove stumps are vertically compressed. This is not due to post-depositional sediment compaction, for compaction after the infilling of the axes would have compacted the silty mudstone outside the axes more than the sandstone inside the axes. This suggests that the vertical compression of the axial systems occurred before being infilled with sand. However, if Unit B was the soil in which these axes were rooted for years prior to the death of the plant (as suggested by Clarey and Tomkins 2016), the axes would not be expected to widen and flatten even when they became weak with rotting out of their interiors. Instead, it appears that the stumps and associated axial systems decomposed outside the constraints of an enclosing soil, and collapsed when the stumps settled into Unit B sediment. Rather than being a soil in which the stumps were rooted, Unit B appears be a subaqueously deposited sandy mud upon which the lycopod stumps settled.
Bioturbation also suggests Unit B was formed immediately prior to Units C and D. The preservation of laminae in Units B through D, as well as the undisturbed surface of Unit A’s megaripples suggest very little to no bioturbation in any of the Fossil Grove sediments. This implies little to no time elapsed between depositional events—i.e., between the deposition of Units A and B, B and C, and C and D. This, in turn, means that Fossil Grove stumps were set in place soon after Unit A was deposited, and soon before the deposition of Unit C. This argues against these trees growing for years atop a pre-Flood (Unit B) surface before being buried by Flood (Unit C) sediments. The bidirectional current (possible tsunami deposit) suggested by Unit A’s megaripples and Unit B’s basal sediments also suggest that Unit A—before the Fossil Grove trees were set in place—was deposited under the kind of catastrophic conditions expected in the Flood.
Contact metamorphism also suggests Unit B was formed immediately prior to Units C and D. Despite the fact that the dolerite is a sill, Fossil Grove sediments show astonishingly little evidence of contact metamorphism. The dolerite itself shows obvious evidence of being intruded into a cold country rock—e.g., chilled margins (Allison and Webster 2017; McCallien 1938; Young and Glen 1888) and chemical alteration supposedly from CO2 released from the intruded shales (Keen 1992; McCallien 1938). However, the intruded sediments are very little altered. Young and Glen (1888) claim metamorphism not only led to “hardening, crushing and burning” of the shale beds, but also rendered plant remains “more obscure” than they otherwise would be. McLean (1973, 30) claims Unit X sediment is “slightly baked.” However, these evidences must have been quite slight, for this author saw no obvious evidence of contact metamorphism in the Fossil Grove sediments. This includes (1) the silty mudstone of Unit B in direct contact with dolerite over the very large surface area of the Fossil Grove floor, (2) the sands of Unit C inside stumps cut through directly by dolerite veins, and (3) xenoliths of Unit X sediment only a few inches thick completely surrounded by dolerite. All this collectively suggests that at the time of the intrusion of the dolerite, the Fossil Grove sediments—especially those of Units B, C, and X—were equally water-soaked (for water to take the heat in lieu of the sediments), and uncompacted and porous (for hot water to be readily transported from the sediments). This makes sense if, in fact, all the Fossil Grove sediments—including Unit B—were rapidly deposited in water and (while still water-logged) were soon thereafter intruded by the dolerite. This is consistent with all the Fossil Grove sediments, including Unit B (older than the stumps), being deposited in Noah’s Flood, and not consistent with Units C–E being Flood sediments and Unit B being a pre-Flood paleosol existing for many years in the pre-Flood world. This, in turn, suggests that the Fossil Grove stumps were settled (allochthonously) into or onto the sediments of Unit B during Noah’s Flood.
Fossil Grove Stumps
Fossil Grove preserves nine stumps with attached axial structures (Stumps, 1, 3, 5–11 in fig. 4), one stump without its axial structure (Stump 2 in fig. 4), one system of axial structures without its stump (Stump 4 in fig. 4), and one extended stigmaria which may or may not be attached to a stump (“Log F” in fig. 4). The original fossils were thin layers of coalified organic material—sloughed off during excavation—surrounding sandstone cores. Before the organic material sloughed off, the sandstone in the cores of each stump was continuous with the sandstone of Unit C. The stump heights range from 4.2 in to 19.2 in (table 1 of Gastaldo 1986; Keen 1992), the sandstone cores of the stumps show vertical lineations (Kidston 1888), the stump diameters range from 13 in to 39 in (table 1 of MacGregor and Walton 1948), and the stumps show lateral compression averaging 0.75:1, with long axes in roughly the NE–SW direction (table 1 of MacGregor and Walton 1948). The axial structures attached to the stumps range from 0 ft to 6 ft in length, bifurcate roughly every 2 ft, exhibit a vertical compressed of about 1:2, and assume a near-horizontal orientation within a foot or so from the stump.
Since the sand of Unit C is continuous with the sand inside the stumps and their axial structures, all investigators (Allison and Webster 2017; Gastaldo 1986; Gunning 1995; Hall, Browne, and Forsyth 1998, 45; Keen 1992; Lawson and Lawson 1976, 72; MacGregor and Walton 1948; Owen et al. 2007; Young and Glen 1888) have concluded that the stumps seen today are actually internal molds of the original stems and axial systems. That is to say, at the time of burial of the upper end of the stumps, Unit C sand infilled hollow stems and axial systems. The vertical lineations on the outside of the stump molds and the bifurcating morphology of the axial systems are characteristic of arborescent lycopods (Cleal and Thomas 1995; Keen 1992; Kidston 1888; Lawson and Lawson 1976, 72; MacGregor and Walton 1948; McLean 1973). However, these characters do not allow genus-level identification, for external bark patterns are used to distinguish lycopod genera (Keen 1992; Kidston 1888; MacGregor and Walton 1948; McLean 1973). Kidston (1888) identified the stumps as Lepidodendron veltheimianum, based on stems and branches found in Unit C (e.g., A, B, C, X in fig. 4) and because that was the most common lycopod species in the Scottish Carboniferous. Although subsequent authors (e.g., Allison and Webster 2017) commonly identify the stumps as Lepidodendron (Keen 1992, 67), and some even claim this is a monospecific stand of Lepidodendron (e.g., Clarey and Tomkins 2016; Gastaldo 1986), identification to the level of the Lepidodendron genus should only be considered probable (as do MacGregor and Walton 1948 and Keen 1992).
After Young and Glen (1888) noted the oval cross section of what was later labeled Stump 11, MacGregor and Walton (table 1 of MacGregor and Walton 1948) extended the observation to all the stumps, and this curious fact has been mentioned by most later writers (Allison and Webster 2017; Cleal and Thomas 1995, 190; Gastaldo 1986; Gunning 1995; Keen 1992; Lawson and Lawson 1976, 71; McLean 1973; Owen et al. 2007), with the major axis of the cross sections in the NE–SW orientation (Gastaldo 1986; Keen 1992; MacGregor and Walton 1948). MacGregor and Walton (1948) believed the deformation was due to tectonic compression of the sediments, McLean (1973) added evidence from a “small fold” in the strata at the western end of the building, and Lawson and Lawson (1976) concurred. Gastaldo (1986), however, observed that the axial systems did not show lateral compression, so, by similarity with lycopod specimens at other localities, suggested that fast-flowing water deformed the trunks while the axial systems of the stumps were buried by (Unit B and lower Unit C) sediment. Gastaldo felt this interpretation was supported by the alignment of the long axes of the stumps corresponding to the alignment of the long axes of prostrate logs and the direction of current suggested by asymmetric ripples—all in Unit C (see fig. 5). All following Gastaldo have accepted Gastaldo’s reinterpretation (e.g., Allison and Webster 2017; Keen 1992; Owen et al. 2007).
Consideration of Unit A’s possible megaripples suggests another interpretation of the environment of deposition of Fossil Grove stumps. The stumps tend to lie in the troughs of the dolerite’s undulations (fig. 5)—thus in the troughs of Unit A’s possible megaripples. Furthermore, the prostrate stems in Unit B (Log A and the lower-placed Log B in fig. 4) have orientations that may be askew of the paleocurrent indicators from Unit C (the long axes of the stumps, the long axes of prostrate stems, and the ripple marks). Log B, lower in Unit B, seems to be very nearly orthogonal to the paleocurrent indicators in Unit C and Log A in the upper part of Unit B, or atop it, seems to lie at an orientation between that of Log B and the paleocurrent indicators in Unit C (see fig. 5). Though the orientation of Log B is out of line with the paleocurrent indicators in Unit C, it is roughly in line with the current direction indicated by Unit A’s possible megaripples. This suggests that Units A and B (along with the original stumps and Logs A and B) were deposited in currents from a NW or SE direction—or bidirectional NW–SE currents, whereas Unit C was deposited in a southwest-trending paleocurrent.
One final consideration of Fossil Grove stumps and lycopod morphology offers further insight into deposition and burial of these specimens. As already mentioned, lycopod stigmaria have appendage scars on all sides. Lycopod stigmaria appendages can also be quite long. Although most fossil specimens show inches of length, appendages of several feet are known. Whether rooted in soil or suspended in water, stigmaria appendages wouldn’t seem to function properly if the stigmaria were too close to horizontal in orientation—i.e., if the stigmaria were too close to the top of the soil if rooted in soil, or too close to the top of the water column if suspended in water. Appendage scars on stigmaria and the length of stigmaria appendages would thus suggest that axial structures in life should have descended rather steeply into whatever medium they grew. Yet, the axial structures of Fossil Grove stumps are all nearly horizontal for up to three axial bifurcations away from the stump. This suggests that these axial structures are not in situ. Rather, the axial structures (structurally weakened by internal decomposition) were apparently bent into a near-horizontal orientation as the stumps settled on some sort of resistant surface. The unnatural bending upward would also explain the flattened nature of these axes. This, of course, would mean that prior to deposition, the axial systems were not in sediment, but suspended in water.
The following depositional model is suggested here to explain the sediments and fossils at Fossil Grove. Very high energy (possibly tsunami) NW–SE current flow deposited Unit A and generated surface megaripples in Unit A—perpendicular to the axis of Midland Valley, so possibly due to earthquake activity on fault(s) currently bounding the Midland Valley to the north and south. As energy waned following the deposition of Unit A, successively less buoyant sedimentary particles settled atop Unit A—first the silty muds of Unit B, then Stumps 1–11 and Logs A and B. The stumps, already decomposed during a lengthy post-death float time, settled onto and into the Unit B surface, bending and flattening the axial structures in the process. Subsequently, a sand-bearing current from the northeast—along the axis of the Midland Valley—brought in sand to finish burying the axial structures of the stumps, then the stumps themselves, deforming the decayed stumps, infilling them and their axial structures, bringing in and orienting hollow trunks and stigmaria, and generating ripple marks on bedding surfaces. Not only would this mean that the stumps were deposited allochthonously, but it would mean that all the sediments of Units A–D and their contained fossils were deposited in the same event—i.e., in Noah’s Flood, if the sediments of C–D are thought to be Flood deposits.
Pre-Fossil Grove Flood Sediments
As part of their argument that Fossil Grove is autochthonous, Clarey and Tomkins (2016) argue that no Paleozoic sediments underlie Unit B. They argue that whereas Silurian, Lower Devonian, Upper Devonian, and earlier Carboniferous sediments are found in other parts of Scotland, they pinch out as Fossil Grove (Glasgow) is approached. The general consensus of United Kingdom geologists (e.g., Bluck 2002) seems to concur with Clarey and Tomkins with respect to the locally deposited, unconformity-bounded stratigraphic sequences of Silurian and Lower Devonian rocks (what used to be called the Lower and Middle Old Red Sandstone).
However, United Kingdom geologists are not in agreement with Clarey and Tompkins’ (2016) claims about the absence of Upper Devonian and earlier Carboniferous sediments under Glasgow. Most geologists put the Upper Devonian and lower Carboniferous sediments together (up through the sediments of Fossil Grove), with just two local to regional unconformities (at the base and top of the Strathclyde volcanics). This is how MacGregor and Walton (fig. VIII of MacGregor and Walton 1948), Hall, Browne, and Forsyth (fig. 2 of Hall, Browne, and Forsyth 1998), and Allison and Webster (fig. F2 of Allison and Webster 2017) represent their stratigraphic columns of the “Glasgow district” or “Glasgow area.” This is also how McLean (fig. 3 of McLean 1973) and Monaghan (fig. 4 of Monaghan 2014) represent it in their cross sections through Glasgow—at least down to and including the Strathclyde volcanics.
The reason the geology is so uniformly interpreted is probably due to the bedrock geology map of the Midland Valley (see figs. 6 and 7, and as seen in geologic maps of fig. 2 of McLean 1973, and fig. 1 of Hall, Browne, and Forsyth 1998, and fig. 9.2 of Read et al. 2002, and figs. 3 and 5 of Monaghan 2014). Surrounding Glasgow to the SW, NW, and SE, and open to the NE, are concentric horseshoe-shaped surface exposures of increasingly older sedimentary units as one travels away from Glasgow. For example, as seen in fig. 7, from Fossil Grove (in the Limestone Coal Formation), traveling to the west, one encounters first the Lower Limestone Formation (which underlies the Limestone Coal Formation), then the sediments of the upper Strathclyde Group (which underlie the Lower Limestone Formation), then the volcanics of the lower Strathclyde Group (which underlie the upper Strathclyde Group), then the sediments of the Inverclyde Group (which underlie the Strathclyde Group), and then the sediments of the Stratheden Group (Upper Devonian sediments which underlie the Inverclyde Group). The same pattern is seen traveling north (with the exception of the absence of Uppermost Devonian sediments) and traveling southeast (after first moving up to the Coal Measures before dropping back down to the Limestone Coal Formation) (fig. 7). This outcrop pattern is that of a NE/SW-oriented syncline plunging to the northeast and nearly centered on Glasgow. This is also consistent with the observed dip of the Fossil Grove strata. If the Limestone Coal Formation (containing the Fossil Grove fossils) is 150 m thick (as interpolated from the sections illustrated in fig. 11 of Hall, Browne, and Forsyth 1998), the distance between upper and lower boundaries of the Limestone Coal Formation in the vicinity of Fossil Grove (see fig. 2) suggest about a 3° dip to the northeast for Glasgow strata. This corresponds to claims of northeast (MacGregor and Walton 1948) and “gentle” northeast dip (Allison and Webster 2017; Gunning 1995; McLean 1973) for the Fossil Grove rocks. The simplest interpretation of the plunging syncline pattern of rocks around Glasgow is to project all of these sedimentary units under Glasgow, and this is exactly what UK geologists have done for many decades. Clarey and Tompkins (2016) argue that Glasgow is located on a topographic high that did not receive sediments until the stratigraphic level of Fossil Grove. This means that the later deformation that created the plunging syncline centered that deformation on the crystalline rocks of a topographic high. Though this is not impossible, this is not likely.
A second problem with claiming no pre-Fossil Grove deposits under Glasgow is how steep the gradient on the Fossil Grove high would have to be to keep all pre-Fossil Grove deposition below the altitude of Fossil Grove (which Clarey and Tomkins seem to think sat close to sea level before the Flood). Devonian sediments outcrop only about 10 mi away from Glasgow to the northwest and 20 mi to the west and south (see fig. 6). Yet the stratigraphic columns of MacGregor and Walton (fig. VIII of MacGregor and Walton 1948), McLean (fig. 1 of McLean 1973), Hall, Browne, and Forsyth (fig. 2 of Hall, Browne, and Forsyth 1998), and Monaghan (table 2 of Monaghan 2014) represent these units with a total thickness of (respectively) over 2700 ft (not representing an unspecified amount of Strathclyde volcanics), 6300–9500 ft, 5500 ft, and 10,090 ft (without the Lower Limestone Formation). This requires that thousands of feet (perhaps as much as nearly 2 mi!) of sediments were deposited and thousands of feet of lava was extruded within 20 mi of Fossil Grove in three compass directions—all the while not disrupting the stumps, roots, and mud of the Fossil Grove forest. This seems very nearly impossible.
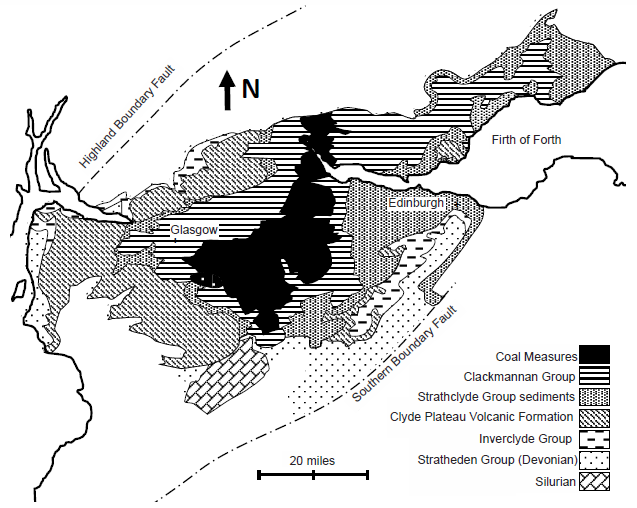
Fig. 6. Geology of the Midland Valley, Scotland. Bedrock geology adapted from fig. 9.2 of Read et al. 2002 and fig. 2 of McLean 1973.
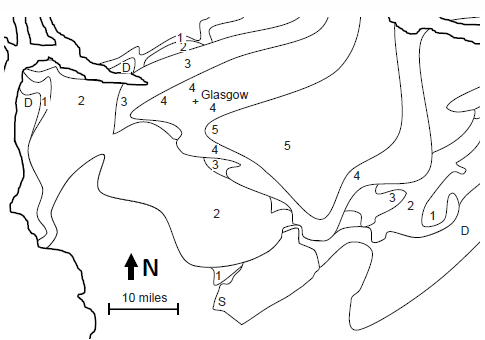
Fig. 7. Simplified geology of the Plunging Syncline of the Glasgow Region. Simplified geology of fig.6. Key: D: Devonian; S: Silurian; 1: Inverclyde Group; 2: Clyde Plateau Volcanic Formation of the lower Strathclyde Group; 3: upper Strathclyde Group sediments; 4: Clackmannan Group; 5: Coal Measures.
A third problem with claiming no pre-Fossil Grove deposits under Glasgow, is how this runs counter to available subsurface data. Although Clarey and Tomkins (2016) rightly point out the absence of boreholes directly under Glasgow, a number of boreholes and surface sections are known within 20 mi of Glasgow in all directions. Although isopach maps constructed from the subsurface data suggest Glasgow sits right on the edge of zero-thickness Inverclyde Group sediments (fig. 9.10 of Read et al. 2002), they also suggest between 1600 ft and 3200 ft (interpolated at about ½ mi) of Strathclyde Group rocks (fig. 9.13 of Read et al. 2002) and 300–650 ft (interpolated at about 525 ft) of Lower Limestone Formation (fig. 9.27 of Read et al. 2002) rocks underlie Glasgow. This is neither an unusually thin expression of these units (roughly half of their thickness at their thickest expression), nor is this a peripheral location for deposition of these units. Rather, Fossil Grove is on the axis of a depositional trough for each unit! For the Fossil Grove to be above deposition while these sediments were deposited, the subsurface data suggest that Fossil Grove would have to lie in the axis of a depositional trough while nearly 2000 ft of sediments thinned to zero thickness on all sides of the Fossil Grove high within the distance of just 10 mi! Again, although such topography is not impossible, it seems quite improbable. What is more likely is that more than ½ mi of Strathclyde sediments and lavas underlie Fossil Grove.
A fifth problem with claiming no pre-Fossil Grove deposits under Glasgow, is how the absence of Lower Limestone Formation under Fossil Grove can be reconciled with the finer-scale geologic maps of Hall, Browne, and Forsyth (fig. 1 of Hall, Browne, and Forsyth 1998) and Allison and Webster (fig. FG2 of Allison and Webster 2017). As was true for Glasgow in the Midland Valley, Fossil Grove in Glasgow is on or near the axis of a syncline plunging to the northeast, with lower-lying stratigraphic units to the south, west, and north and overlying stratigraphic units to the southeast, east, north, and northwest (see fig. 2). This, again, suggests that the underlying Lower Limestone Formation—exposed at the surface less than half a mile to the southwest of Fossil Grove—actually underlies Fossil Grove. Furthermore, the surface expression of the sill located at Fossil Grove (following the bedding of the Fossil Grove sediments) mirrors the surface expression of the sill located in the Lower Limestone Formation a half mile to the southwest, as well as the surface expression of ironstone bands and coal seams in the Limestone Coal Formation sediments overlying the Fossil Grove sediments (see fig. 2). This suggests that the sediments of the Lower Limestone and Limestone Coal Groups have been deformed together, and probably deposited together (for later igneous sills to insert themselves between sedimentary layers in the same manner). Additionally, the surface expression of the Lower Limestone and the Limestone Coal Formations in the Fossil Grove vicinity suggests the former is about half the thickness of the latter, consistent with the relative thicknesses deduced from isopachs of subsurface data (~525 ft of Lower Limestone from fig. 9.27 of Read et al. 2002 and ~1115 ft of Limestone Coal Formation from fig. 9.28 of Read et al. 2002). This seems to offer further credibility to the isopach modelling and incorporates surface data located within 2 mi of Fossil Grove. It also suggests that there is probably something on the order of 525 ft of Lower Limestone Formation sediments beneath Fossil Grove.
A sixth problem with claiming no pre-Fossil Grove deposits under Glasgow, is how the absence of Limestone Coal Group sediments beneath Fossil Grove can be reconciled with the very fine-scale geologic map of Allison and Webster (fig. FG2 of Allison and Webster 2017). Very near the axis of the plunging syncline running just west of Glasgow, Fossil Grove lies about 30% of the way between the contacts of the Limestone Coal Formation with the Lower Limestone and Upper Limestone Formations, and about halfway between the base of the Limestone Coal Formation and a set of closely spaced ironstone bands and coal seams of the Limestone Coal Formation (see fig. 2). Hall (fig. 11 of Hall, Browne, and Forsyth 1998) represents two stratigraphic sections of the Limestone Coal Formation—one, the Linwood section, located about 6 mi to the WSW of Fossil Grove and the other, the Colston Road borehole, located about 3.7 mi to the ENE of Fossil Grove (see fig. 2). In both sections the closely spaced ironstone bands and coal seams are in the upper half of the Limestone Coal Formation, placing the Fossil Grove sediments in the lower part of the Limestone Coal Formation, about 30% up the formation from its base. This would be just above the well-known Johnstone Clayband Ironstone of the lower Limestone Coal Formation, the very stratigraphic position that Hall, Browne, and Forsyth (1998, 45) suggested for Fossil Grove. Extrapolating from the Limestone Coal Formation thickness at the Linwood section (410 ft) and the Colston Road borehole (580 ft), the Limestone Coal Formation at Fossil Grove should be about 490 ft thick, placing Fossil Grove roughly 150 ft from the base of the Limestone Coal Formation. This suggests that 150 ft of Limestone Coal Formation sediments underlie the Fossil Grove sediments.
All data considered, it is extremely unlikely that Fossil Grove sediments sit on top of pre-Flood bedrock undisturbed by Flood catastrophism as suggested by Clarey and Tomkins (2016). What is most likely is that more than 1500 ft (and probably closer to 3000 ft) of flood basalts and more than 1700 ft of Flood sediments were deposited during the Flood before the Fossil Grove sediments were deposited. The sub-Fossil Grove geology strongly suggests that the Fossil Grove stumps were deposited allochthonously in the midst of the Flood.
Autochthony Criteria
Clarey and Tomkins (2016) offered seven criteria for identifying autochthonous fossil forests. Given their claim that Fossil Grove was autochthonous based on these criteria, and the strong likelihood that Fossil Grove is not autochthonous, Clarey and Tomkins’ autochthony criteria are unreliable. The criteria given were as follows: (1) “Finding multiple, single-species trees spaced in growth position in the same horizontal plane, nearly equidistantly spaced in all directions from the trunks, tree to tree, and not merely randomly spaced. . . .”; (2) “Finding multiple trees in the same rock layer or along a common surface and/or stigmarian (root) axes interlacing with other trees over a common, three-dimensional, single rock layer. . . .”; (3) “Finding trees with root systems (stigmarian axes) that crosscut bedding layers up to and even exceeding 30° and possessing perpendicularly inserted radiating appendages (rootlets). . . .”; (4) “Evidence of rapid burial by thick sediment and water, which is common for most standing lycopsids. . . ., and multiple trees at a single site that were distorted by water flow in the same direction during burial. . . .”; (5) “Finding a lack of sedimentary rock layers underneath the trees. The presence of Flood-related layers underneath the trees would eliminate the possibility of in situ [sic] growth. In contrast, a lack of sedimentary layers under the trees, other than the fossil soil (paleosol) layer that contains the trees, would support the interpretation of an original pre-Flood setting. In other words, in situ [sic] trees should have no substantial sedimentary layers beneath or fossils beneath or coal layers beneath.”; (6) “Finding no bowing or distortion of any sedimentary layers beneath the tree stumps, which would indicate the tree was transported and emplaced from above during burial, distorting the underlying sediments. . . .”; and (7) “Finding accompanying vegetation like Calamites, although rare, that crosscut the same layers as the lycopod tree stumps . . . .”.
Regarding Clarey and Tomkins’ criterion number one, although it is true that individuals of a given tree species are not distributed randomly, they are also not distributed equidistantly as they claim. In general, pretty much independent of taxon and environment, individuals of sedentary species tend to assume a spatial distribution which is much more clustered than would be expected with either a uniform/equidistant or random pattern (partly because offspring tend to establish themselves close to parents). Also, this criterion must be demonstrated statistically, as it is often very difficult to discriminate between clustered, random, and uniform spatial distributions.17 Consequently, generalizing this criterion beyond the forest ecosystem, an autochthony criterion should read something like: “Individuals of sedentary species are spaced in the non-random, clumped pattern characteristic of living organisms.”
Regarding Clarey and Tomkins’ criterion number two, most of those who have speculated that arborescent lycopods grew floating in water, have also speculated that multiple individuals interlaced their stigmarian axes so as to maintain their trunks in a vertical position (e.g., Bruce 2002; Kuntz 1884, 1895; Scheven 1981; Wieland 1995). If this is so, the interlacing may have been sufficient to keep adjacent trees together in that interlaced condition even after the lycopod forest was torn apart by the catastrophic conditions of the Flood. If individual arborescent lycopod stumps could float in an upright position, it seems just as possible that multiple arborescent lycopod stumps could have floated together with their stigmarian axes interlaced. Thus, Gastaldo’s suggestion of interlaced stigmarian axes is not a definitive criterion of autochthony. On the other hand, establishing that a rock layer in which multiple individuals are found is actually a paleosol or hardground would be good evidence of autochthony.18 Consequently, generalizing this criterion beyond the lycopod forest, dropping non-definitive criteria, and including definitive criteria, an autochthony criterion should read something like: “Multiple individuals are in growth position in the same paleosol or attached to the same hardground.”
Regarding Clarey and Tomkins’ criterion number three, because the arborescent lycopod stigmarian appendages are rather long and exit perpendicular to the stigmaria around the whole stigmarian circumference, lycopod axial structures must have departed the trunk at a steep angle to the horizontal until they had reached at least a couple feet beneath the surface. If these stigmarian axes were normally suspended in water and if the stump with attached axial structures could float in growth-like orientation, then its axial structures could get buried at a high angle with respect to the enclosing sediment—even if they were deposited allochthonously. Thus stigmaria and/or stigmarian appendages oriented at an angle with respect to bedding does not seem to be a conclusive criterion. Furthermore, the fact that stigmaria can maintain their three-dimensional form even when hollowed out by decomposition suggests that the outer surface of stigmaria—like lycopod “bark”—is rather stiff. This stiffness may allow even hollowed stigmaria to get buried across sedimentary bedding. In fact, this author’s experience with Carboniferous strata suggests that even flexible allochthonous structures can crosscut strata. For example, stigmarian appendices (“rootlets”) are often found curving or even drooping as if they were flexible in life, especially those that are still attached to stigmaria. Appendices that exit the stigmaria in the plane of bedding tend to follow the plane of bedding, whereas those that exit the stigmaria at successively higher angles to the planes of bedding are successively more bent towards the planes of bedding, so that long appendages all end up parallel to the plane of bedding. This bending is not consistent with simple compression of the sediment and seems to be indicative of free-floating appendages being buried in soft sediment for the first time at the time of burial. Thus I would consider this pattern of stigmaria/appendage orientations indicative of allochthony. On the other hand, those very specimens indicate that even (flexible) appendages can be oriented at high angles to the plane of bedding. Notwithstanding all that has been said about stigmaria and their appendages in particular, growth orientations which are contrary to likely transport orientations could be definitive evidence of autochthony. Consequently, generalizing this criterion beyond the lycopod forest, dropping non-definitive criteria, and including definitive criteria, an autochthony criterion should read something like “Organisms are in growth orientation, especially when that orientation is contrary to expected orientation of transport and/or deposition.”
Regarding Clarey and Tomkins’ criterion number four, paleocurrent deformation of vertical trunks without a corresponding deformation of stigmarian structures only argues that when the vertical trunks were being deformed, the stigmarian structures were already buried. If, in fact, arborescent lycopod stumps did float in growth orientation, then burial would bury the stigmarian structures first. Thus even under allochthonous conditions lycopod trunks could get deformed by currents without a corresponding deformation of the stigmarian structures. Unlike Clarey and Tomkins’ (2016) claim, the paleocurrent deformation of Fossil Grove trunks without a corresponding deformation of stigmarian structures is not definitive evidence of autochthony. Furthermore, it seems possible to explain lateral deformation of hollow trunks by current flow from two different directions—either current flow that extends the downstream side of the trunk in the direction of flow (what Gastaldo suggests for the Fossil Grove stumps) or current flow that collapses the upstream side of the trunk towards the downstream side of the trunk. In fact, in Gastaldo’s model it is not clear why the current would not also bend the upstream side of the trunk, creating a u-shaped trunk cross section, open on the upstream side. Given that each model results from paleocurrents in opposite directions, and the two models suggest paleocurrents in orthogonal directions, a given set of trunk deformations could be argued consistent with four different paleocurrent directions. Given the real world’s cloud of data points around expected values, it is not clear that this is a good paleocurrent indicator at all. Regarding rapid burial in general, rapid burial might well be expected in the case of Flood burial of in situ pre-Flood communities. However, preservation of pre-Flood communities in pre-Flood times would not necessarily occur under catastrophic conditions. Consequently, non-catastrophic burial (in pre-Flood times) might occur in the case of in situ communities and catastrophic burial (during the Flood) might occur in the case of allochthonous communities. It would seem that catastrophic burial is an indicator of neither allochthony nor autochthony. However, most of the substrates of pre-Flood communities were probably not put in place catastrophically. Therefore, the substrates of in situ pre-Flood communities should show evidence of slow development. Consequently, correcting this criterion, extending it beyond the lycopod forest, and dropping non-definitive criteria, an autochthony criterion should read something like: “Evidence should exist for the slow development of substrate in autochthonous communities.”
Regarding Clarey and Tomkins’ criterion number five, given the possibility of the formation of pre-Flood sediment and fossils, this criterion needs to be worded carefully. Consequently, generalizing this criterion beyond a forest, an autochthony criterion should read something like: “There are no underlying Flood sediments.”19
Regarding Clarey and Tomkins’ criterion number six, if something is being deposited allochthonously, it is usually being deposited in place with a variety of other sedimentary particles of similar hydrodynamic properties. Most of the time this should result in no to very little density difference between the transported trees and the surrounding sediment, and thus no differential settling forces on the underlying sediment. Given, then, that the bowing of underlying sediment is not the usual expectation of allochthony, the absence of bowing in underlying sediment is not discriminatory between allochthony and autochthony. This is in spite of the fact that positive evidence of the bowing of underlying sediments—such as claimed by Oard and Giesecke (2007, 238)—does seem to favor allochthony rather than autochthony. This author suggests generalizing the criterion and including the bowing of underlying sediment as one way to determine if the substrate is as compact, settled, or lithified as it should be in autochthonously formed deposits. In the case of a pre-Flood community being buried in Flood strata, the pre-Flood substrate should be considerably more compacted and settled than the Flood sediments laid down on top of it. In contrast, Flood sediments at the time of deposition should be more porous and deformable than the pre-Flood substrate. Autochthonous assemblages, then, should be found in sediments considerably more compacted, settled, and lithified at the time of burial than the sediment lying atop them. Consequently, generalizing this criterion beyond a forest, an autochthony criterion should read something like “The substrate for the autochthonous community should be much more compacted, settled, and/or lithified than the sediment atop it at the time of burial.”
Regarding Clarey and Tomkins’ criterion number seven, in principle, this criterion is one example of a more general one (“Organisms are in growth orientation, especially when that orientation is contrary to expected orientation of transport and/or deposition”), so it does not warrant the status of a separate criterion. Furthermore, on the particular consideration of Calamites, as Fayol (1886) and Coffin (1971, 1975) discovered in flotation experiments, Equisetum—the closest living equivalent to the extinct Calamites—floats vertically in water, either in clusters attached by roots, or as individual stems. Furthermore, the tentaculitoid tubeworms (“Spirorbis” in old literature) not uncommonly attached to vertically oriented Calamites stems (e.g., at Joggins, Nova Scotia, as illustrated in fig. 17 of Dawson 1855; Lyell and Dawson 1853), may be marine organisms20 (Coffin 1968). If so, they are usually interpreted to indicate that Calamites floated vertically in the marine realm for some time prior to burial—and maintained that vertical orientation during burial. Thus, because vertical is an expected orientation of transport of Calamites, vertical Calamites is not a good indication of autochthony.
To add to these criteria, this author suggests the following: (1) On a spectrum of disparity, biological communities are typically highly disparate (e.g., all the kingdoms of bacteria, algae, protists, fungi, animals, plants and multiple classes, orders, families, and genera in each category). In contrast, sorting (e.g., by Flood waters) tends to separate disparate objects into monotypic assemblages. Autochthonous assemblages, then, should display a very much higher disparity of higher taxa than will be displayed in allochthonous assemblages.21 (2) On a spectrum of niche diversity, biological communities typically display high niche diversity (producers and consumers and decomposers, herbaceous and woody plants, tall and short trees, tall and short bushes, etc.). In contrast, sorting (e.g., by Flood waters) tends to separate objects from different niches. Autochthonous assemblages, then, should display a very much higher disparity of niches than will be displayed in allochthonous assemblages. (3) With regard to age distribution, biological communities typically display a full age distribution for each species (juvenile, young, middle-aged, aged). In contrast, because sorting tends to separate specimens by size, sorting by Flood waters might be expected to separate organisms of different ages, and sometimes even gender. Autochthonous assemblages, then, should display more of the usual age distribution of a population than will be displayed in allochthonous assemblages. (4) Regarding the diversity of plant or animal parts, communities typically display a full range of plant and animal parts at a given time (rootlets and roots and trunks and branches and stems and leaves; heads, arms, legs, trunks, etc.). In contrast, sorting (e.g., by Flood waters) tends to separate different plant and animal parts. Autochthonous assemblages, then, should display a more complete range of plant and animal parts than will be displayed in allochthonous assemblages. (5) Infaunal organisms are so abundant and active in modern communities that the sediment substrates of modern communities are heavily bioturbated, usually completely obliterating original bedding. In contrast, the Flood usually involved a sediment production rate that was too high to permit extensive infaunal bioturbation (e.g. Brand and Chadwick 2016, 1086–1097). Autochthonous assemblages, then, should display much more substrate bioturbation than will be displayed in allochthonous assemblages. (6) In general, the more rapid the burial of a community, the more likely the resultant fossils are in situ. The longer the time elapsing from the death of the community to the community’s burial, the more likely that transport will have moved the individuals of the community to some place other than where they lived. Autochthonous assemblages should evidence much less pre-burial organism decomposition than is evidenced in allochthonous assemblages.
Arranging these additional criteria and the revisions of Clarey and Tomkins’ criteria this author suggests the following 12 criteria would be more appropriate for identifying in situ pre-Flood communities:
Autochthonous fossil assemblages should
- not lie atop Flood sediments [vs. lie atop Flood sediments].
- include a very high disparity of higher taxa [vs. monotaxic].
- include a very high disparity of niches [vs. monotaxic].
- include the usual age distribution of member populations [vs. one age bracket].
- include the full range of plant and animal parts found in living organisms [vs. one plant or animal part].
- contain sedentary individuals attached to the same paleosol or hardground [vs. unattached, in different layers, no evidence of paleosol or hardground development].
- include a thoroughly bioturbated substrate [vs. minor bioturbation].
- contain a much more compacted, settled, and lithified substrate than the sediment atop them at the time of burial [vs. the same degree of compaction, porosity, and lithification].
- contain sedentary individuals spaced in the non-random, clumped pattern characteristic of living organisms [vs. alignment or randomness].
- contain organisms in growth orientation, especially when that orientation is contrary to expected orientation(s) of transport and/or deposition [vs. in non-growth depositional orientation].
- evidence little to no pre-burial organism decomposition [vs. much pre-burial decomposition].
- evidence slow development of both substrate and burial sediments [vs. catastrophic development of both].
The closer a given fossil assemblage is to what is expected of autochthonous communities on the spectrum between that and what would be expected with Flood-related allochthony (what is included in the square brackets), the less likely the assemblage has an allochthonous origin in the Flood.
Table 1. Fossil Grove Autochthony vs. Allochthony.
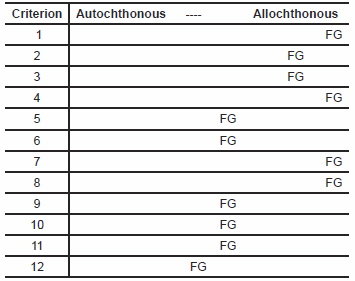
Let us now consider Fossil Grove in the light of our twelve criteria. First, it appears as if there is something in excess of 4000 ft of Flood strata beneath Fossil Grove, including more than 1700 ft of Flood sediment. This places Fossil Grove definitively at the allochthonous end of the spectrum (table 1, line 1). Second, other than Arenicola annelid burrows on the bedding planes of Unit C sandstones (Young and Glen 1888)—and thus from a different community—all Fossil Grove fossils are attributable to the Order Lycopodales, and likely all to the genus Lepidodendron, and maybe even a single species in that genus (Kidston 1888 assigns them all to Lepidodendron veltheimianum). The monospecific—or nearly monospecific—nature of Fossil Grove is most consistent with allochthony (table 1, line 2). Likewise (and third), other than, again, fossil worm traces from another community, Fossil Grove fossils are from a single (tall tree) forest niche. This again places Fossil Grove solidly in the allochthonous end of the spectrum (table 1, line 3). Fourth, the age-datable (by size) lycopods at Fossil Grove are found on two horizons—11 large-diameter stumps, one large-diameter Log B and one medium-diameter Log A in Unit B and/or lower Unit C, and several medium-to-small–diameter logs in the middle or upper Unit C. Fossil Grove’s restricted age distribution, combined with the possible separation of specimens into different units according to size is strongly suggestive of allochthony (table 1, line 4). Fifth, Fossil Grove lycopods are represented by two of the 4–6 logs, all 11 stumps, all but one of the stigmaria, and an unspecified number of stigmarian appendages in Unit B, and 2–4 of the 4–6 logs, a single stigmarian fragment, and rare leaf fragments in Unit C. Although there is some diversity of plant parts, the evidence that most plant parts are separated into different layers tips the evidence in the direction of allochthony (table 1, line 5). Sixth, although the bases of the Fossil Grove stumps are found in a single stratum (Unit B), there is no evidence of paleosol development in that stratum. This might be judged partial evidence of allochthony (table 1, line 6). Seventh, the preservation of original depositional laminae in Unit B and the corresponding lack of bioturbation is most consistent with allochthony (table 1, line 7). Eighth, the mild- to non-baked nature of Units B, C, and X, even in direct contact with dolerite sills, suggests Units A and B have the same porosity and compaction as Units C and X at the time of the intrusion of the sill. If the sill and Units C and X were formed in the Flood, then Units A and B were as well, and the Fossil Grove fossils would have to be allochthonous. Thus the compaction and porosity of Fossil Grove deposits indicates allochthony (table 1, line 8). Ninth, Fossil Grove stumps appear to be spatially positioned in the troughs of what might be megaripples in Unit A. If this pattern is real, this type of alignment is strongly suggestive of allochthony (table 1, line 9). Tenth, the horizontal orientation of the axial structures of Fossil Grove stumps may not be growth orientation. Deformation of what were originally more steeply dipping axial systems would also better explain the vertical compression of those axial structures than the collapse of axial structures in the original rooting substrate. The non-growth orientation of Fossil Grove stumps is thus strongly suggestive of allochthony (table 1, line 10). Eleventh, the fact that Fossil Grove stumps are internal sand molds of stumps and associated axial systems, and that the remaining stigmaria, trunks, and branches are either collapsed or filled with sand, suggests that all the Fossil Grove lycopods had experienced considerable decay long before burial. Although it is possible for a forest to be destroyed and experience decomposition before burial, for this to occur without evidence of community regeneration seems unlikely. Thus the highly decomposed state of Fossil Grove plants is more suggestive of allochthonous burial than autochthonous (table 1, line 11). Twelfth and finally, all the strata of Fossil Grove have evidence of rapid deposition, including Unit A (with its possible megaripples) deposited before the stumps. The catastrophic origin of strata underlying Fossil Grove stumps strongly suggests their allochthonous origin (table 1, line 12). In short, applying the 12 autochthony/allochthony criteria to the Fossil Grove site leads unequivocally to Fossil Grove being allochthonous in origin.
Paleozoic Standing Forests in General
Of all the claims of in situ coal forests, Fossil Grove is perhaps the most celebrated. But it is far from the only such claim. In fact, DiMichele and Falcon-Lang (2011) not only claim that in situ fossil plants are “common” (p. 585) and “numerous” (p. 602), they also “select” for their table 1 (pp. 588–589), 63 fossil assemblages of multiple in situ plants from the Carboniferous alone, and four more from the lowermost Permian. Since every in situ claim is a challenge to the Flood model, the 12 autochthony/allochthony criteria suggested above should be systematically applied to these claims. Yet, hints that all these claims would all be found to be allochthonous can even be found in DiMichele and Falcon-Lang’s (2011) review. Eleven lycopod stumps were allochthonously deposited in an upright position at Fossil Grove during the Flood. It seems likely, then, that the same Flood would have deposited clusters of lycopod stumps in upright position at many other locations at roughly the same stratigraphic level. According to DiMichele and Falcon-Lang (2011, 585), claims of in situ forests are “far more often” reported from the Pennsylvanian “than in any other interval in the geological record.” And 46 of the 63 Pennsylvanian fossil forests in DiMichele and Falcon-Lang’s (2011) table 1 contain upright lycopod trees. A tendency on the part of arborescent lycopods to float upright during the Flood might not only explain most of DiMichele and Falcon-Lang’s claims of upright fossil forests, but also explain why the Pennsylvanian has more examples than any other part of the fossil record.
DiMichele and Falcon-Lang (2011) provide insufficient information in their review to apply most of the 12 autochthony/allochthony criteria to the 67 claims of in situ forests of their table 1. However, they provide sufficient information to comment on at least a third of the criteria—the first and last two. Regarding the last criterion, for example, DiMichele and Falcon-Lang (2011) indicate that all in situ fossil forests are not only rapidly buried, but remain under conditions of high sedimentation rate long after burial (so as not to be eroded away). This is consistent with the prolonged rapid deposition expected in a global Flood. Although DiMichele and Falcon-Lang do not discuss the mode of formation of the claimed substrate sediments, a clue about their formation—and an application of Criterion 1—is given by another of their claims. DiMichele and Falcon-Lang (2011) comment that “Many buried Carboniferous forests with tall, upright trees occur above coal beds” (p. 593) (e.g., the Killingwood Colliery claim of 1830 figured in fig. 2 of DiMichele and Falcon-Lang 2011). Given the likelihood that Carboniferous coal beds were themselves allochthonous Flood deposits, this automatically places “many” of the Carboniferous fossil forests overlying Flood deposits—strongly implying allochthonous origin of the fossil forests themselves. Regarding the second criterion of autochthony, 40 of DiMichele and Falcon-Lang’s 67 fossil forest claims in table 1 contain trees from only one higher taxon. This, again, is indicative of allochthony. Regarding the second to the last criterion, 61 of DiMichele and Falcon-Lang’s 67 fossil forest claims in table 1 are preserved as molds—internal molds of hollowed trees. This means nearly all these fossil forests were substantially decomposed at the time of burial—having been deposited long after the trees had died. Such a long time between the death and burial of the trees makes autochthony unlikely. Furthermore, the same unusual taphonomy in such a high percentage of fossil forests suggests a common cause—e.g., a Flood. Interesting enough, of the six fossil forest claims from DiMichele and Falcon-Lang’s table 1 having another mode of preservation, all six contain only one higher taxon in standing position.
Further comments from DiMichele and Falcon-Lang (2011) that may be relevant to autochthony/allochthony criteria are (1) medullosan pteridosperms leaned on one another for support (pp. 585, 597, 600) (which may, instead, be evidence of allochthonous non-growth-orientation of these pteridosperms), (2) taxonomic “heterogeneity . . . appearing stochastic at small spatial scales but with clear gradients at larger scales” (p. 596) (which data may, instead, be explained by sorting of plant parts), (3) spacing of stumps indicate “very open” forest canopies compared to modern rain forests (p. 596) (which data may, instead, be explained as an unnatural spatial distribution of stumps), (4) “it remains puzzling that so few climbers have been unequivocally documented attached to the dominant erect lycopsid trees” (p. 597) (which may be due to separation of plant parts by catastrophism and water sorting), and (5) “A common preservational mode . . . is . . . burial . . . in tidal sediments” (which in conventional theory suggests marine deposition, and may still require marine deposition after creationist reinterpretation). Although a systematic study of in situ fossil forest claims will certainly include some challenges for creationists (e.g., in situ ash-fall claims, root regeneration from vertical Calamites and tree-fern trunks, “tidal” rhythmites), the consilience of clues from DiMichele and Falcon-Lang (2011) suggest that all Carboniferous fossil forests are allochthonous Flood deposits.
Discussion and Conclusion
Multiple evidences (low taxonomic and niche disparity, limited range of plant ages and plant parts, the absence of bioturbation and paleosol development, unnatural horizontality of axial structures, correlation of stump position with possible underlying megaripples, rapid development of all the sediments above and below the stumps, and substantial decomposition of stumps prior to burial) collectively point quite definitively to an allochthonous origin for the stumps at Fossil Grove in Victoria Park in Glasgow, Scotland. Borehole data combined with the bedrock surface pattern from the Glasgow region indicate that Fossil Grove is underlain by at least 1700 ft of Flood sediments and nearly 3000 ft of Flood basalts. Fossil Grove is an example of allochthonous deposition well into the Flood, and not (as suggested by Clarey and Tomkins 2016) an example of an autochthonous forest overwhelmed by the first Flood waters to reach it.
Fossil Grove has never been studied with the kind of detail that the site deserves. The initial publication (Young and Glen 1888), typical of paleontological studies of that time, did not examine the site with the detail common in modern paleontological studies. The next substantial study (MacGregor and Walton 1948)—and, in fact, also the most recent publication (Allison and Webster 2017)—were presented as guidebooks for the general reader, and so do not examine the site with the rigor expected in primary scientific literature. The most careful scientific study of the site (Gastaldo 1986) focused on why the lycopod stumps were oval-shaped in cross section, so did not offer a thorough study of all the site’s data. This paper’s author could examine the site for only a very short time and under extremely poor light conditions. Yet, the important data that arose from each of these studies suggests there is much more to learn from Fossil Grove in a properly thorough study. In fact, such a study of Fossil Grove is expected to reveal considerable data explained adequately only from a young-age creationist perspective. This includes megaripples which may be consistent only with Flood-scale tsunamis, stump decomposition which may be consistent only with the long float times afforded by a months-long diluvial catastrophe, spatial distribution of stumps which may be consistent only with off-shore deposition in the midst of a coastal pounding of the scale of the Flood, rapidly deposited thick sedimentary sequences which may be consistent only with the global-scale deposition of the Flood, biome-wide sorting which may be consistent only with a global deluge, and rapid transport of igneous intrusion heat which may be consistent only with water mobilization on the scale of a global Flood.
Clarey and Tomkins’ (2016) allochthony criteria led Clarey and Tomkins to the incorrect conclusion that Fossil Grove was an autochthonous deposit. A revision and expansion of Clarey and Tomkins’ criteria generates a dozen autochthony/allochthony criteria which, when applied to Fossil Grove, firmly (and correctly) identify Fossil Grove as an allochthonous deposit. DiMichele and Falcon-Lang’s review of what they think are Pennsylvanian in situ fossil forests suggests that application of the 12 criteria to all Carboniferous forests might well conclude they are all allochthonous Flood deposits. This sort of restudy would transfer a number of fossil sites from evidence against a global Flood, to evidence against the conventional understanding of Carboniferous sediments and fossils. Rejecting conventional paleosol claims, this sort of restudy would demand a reevaluation of paleosols in general. Accepting catastrophism for substrate sediments challenges conventional models for Carboniferous environments. Including substrates into the same depositional catastrophes with their overlying sediments expands catastrophism beyond the restricted local scale currently accepted in conventional geology. Recognizing restricted diversity as a consequence of sorting, suggests a linking of widely spaced fossil sites into single diluvial events. For these reasons, and for probably many others besides, a systematic creationist study of fossil forests would generate substantial difficulties for conventional geological models.
Such a systematic study is likely to reveal other indicators of allochthony. A reviewer of an early version of this paper noted that if the allochthonous theory was true, the axial structures of stumps might be deformed and/or oriented by currents during deposition. This author suspects that the axial structures near to the trunk (within 10 ft) were probably too stiff to be deformed by currents, but that axial structures at a greater distance might show orientation by current. Furthermore, isolated stumps (lacking appendages intertwined with other stumps) might be expected to have their overall axial structure pattern oriented with the current. Although this author believes that the Fossil Grove axial structures were too close together to allow independent alignment with current, examination of the axial structures of isolated trees at other localities may demonstrate such current orientation (and thus further evidence of allochthony). The same reviewer asked what happens to the axial structures distally, as this may yield even more data. Unfortunately, the Fossil Grove axial structures terminate without clarifying what happens to the axial structures distally, but examination of fossils at other sites may yield even more allochthony evidence (e.g., that some distal axial structures curve upward).
Clarey and Tomkins (2016) offered the Fossil Grove site as a falsification of floating forest theory. And, if the Fossil Grove trees had been rooted in the soil in the uppermost portion of a regolith atop crystalline basement rock, there might have been reason to believe that at least one of the pre-Flood lycopod forests did not float, but grew on land. However, Fossil Grove was an allochthonously deposited Flood deposit, and it is likely that all claimed in situ Carboniferous fossil forests were actually emplaced allochthonously in the Flood. The allochthonous nature of Fossil Grove suggests that Fossil Grove lycopods grew somewhere else, and suggests that arborescent lycopod stumps floated in an upright orientation. The fact that Fossil Grove lycopods were hollow at the time of burial (as probably was true for most arborescent lycopods) suggests they died long before burial, and thus floated for a very long time (and probably distance) from their original growth positions. The Fossil Grove sediments are probably located less than a dozen stratigraphic feet of an ironstone band with marine fossils, and Fossil Grove’s sandstones contain cross-bedding which may turn out to be consistent only with marine sedimentation. Thus, as is the case with most Paleozoic plants, Fossil Grove lycopods are buried in and/or near marine sediments. This, combined with the absence of land plant morphologies (e.g., true roots), modern land plant taxa, and large, true land animals until after the Carboniferous, suggests the Carboniferous plants are probably part of a marine ecosystem, not a land ecosystem. In fact, examining many sorting-impoverished fossil assemblages (like Fossil Grove) to reconstruct the original ecosystem(s) suggests very complex ecosystems in a large forest biome. Far from being a falsification of floating forest theory, Fossil Grove offers multiple evidences in support of a huge forest biome floating atop the world’s pre-Flood oceans.
Acknowledgements
Although the author takes full responsibility for the inadequacies of the paper as it stands, the author is indebted to the comments of three reviewers for substantial improvements in the paper from an earlier version.
References
Allison, Ian, and David Webster. 2017. A Geological Guide to the Fossil Grove, Glasgow. Glasgow: Geological Society of Glasgow. https://www.geologyglasgow.org.uk/docs/017_070__fossilgrove_1425481004.pdf.
Bluck, B. J. 2002. “The Midland Valley Terrane.” In The Geology of Scotland, 4th edition, edited by Nigel H. Trewin, 149–166. London, United Kingdom: The Geological Society.
Brand, Leonard, and Arthur Chadwick. 2016, Faith, Reason, and Earth History: A Paradigm of Earth and Biological Origins by Intelligent Design, 3rd edition. Berrien Springs, Michigan: Andrews University Press.
Bruce, Wesley. 2002. “The Salinity of a Floating Forest.” TJ 16, no. 3 (December): 95–97.
Clarey, Timothy L., and Jeffrey P. Tomkins 2016. “An Investigation into an In Situ Lycopod Forest Site and Structural Anatomy Invalidates the Floating-Forest Hypothesis.” Creation Research Society Quarterly 53 (Fall): 110–122.
Cleal, Christopher J., and Barry A. Thomas 1995. Palaeozoic Palaeobotany of Great Britain. Geological Conservation Review Series No. 9. London, United Kingdom: Chapman and Hall.
Coffin, Harold G. 1968. “A Paleoecological Misinterpretation.” Creation Research Society Quarterly 5, no. 2 (September): 85–87.
Coffin, Harold G. 1971. “Vertical Flotation of Horsetails (Equisetum): Geological Implications.” Geological Society of America Bulletin 82, no. 7 (July): 2019–2022.
Coffin, Harold G. 1975. “Research on the Classic Joggins Petrified Trees.” Creation Research Society Quarterly 6, no. 1 (June): 35–44, 70.
Dawson, John William. 1855. Acadian Geology: An Account of the Geological Structure and Mineral Resources of Nova Scotia, and Portions of the Neighbouring Provinces of British America. Edinburgh, United Kingdom: Oliver and Boyd.
DiMichele, William A., and Howard J. Falcon-Lang. 2011. “Pennsylvanian ‘Fossil Forests’ in Growth Position (T0 Assemblages): Origin, Taphonomic Bias and Palaeontological Insights.” Journal of the Geological Society, London 168 (February): 585–605.
Fayol, Henri. 1886. Études sur le Terrain Houiller de Commentry. Livre Premier: Lithologie et Stratigraphie. Bullètin de la Sociéty de l’Industrie Minérale, Ser. 2, Vol. 15, no. 3–4: 1–543.
Gastaldo, Robert A. 1986. “An Explanation for Lycopod Configuration, ‘Fossil Grove’ Victoria Park, Glasgow.” Scottish Journal of Geology 22, no. 1 (May): 77–83.
Glasgow City Council. n.d. “Victoria Park Heritage Trail”. Glasgow, United Kingdom: Glasgow City Council. https://www.glasgow.gov.uk/CHttpHandler.ashx?id=31483&p=0.
Gunning, Alastair. 1995. The Fossil Grove. Glasgow, United Kingdom: Glasgow Museums.
Hall, I. H. S., M. A. E. Browne and I.H. Forsyth. 1998. Geology of the Glasgow District: Memoir for 1:50000 Geological Sheet 30E (Scotland). British Geological Survey Memoir. London, United Kingdom: British Geological Survey.
Keen, M. C. 1992. Excursion 2: Fossil Grove. In Geological Excursions Around Glasgow and Girvan, edited by James D. Lawson, and D. S. Weeden, 63–70. Glasgow, United Kingdom: The Geological Society of Glasgow.
Kidston, Robert. 1888. “Note on the Nature of the Fossil Trees Found at Whiteinch.” Transactions of the Geological Society of Glasgow 8, no. 2: 235–236.
Kuntze, Otto. 1884. Phytogeogenesis: Die Vorweltcliche Entwicklung der Erdkruste und der Pflanzen, Phytogeogenesis. Leipzig, Germany: Paul Frohberg.
Kuntze, Otto. 1895. Geogenetische Beiträge. Leipzig, Germany: Gressner and Schramm.
Lawson, Judith Ann and James D. Lawson. 1976. Geology Explained around Glasgow and Southwest Scotland, Including Arran. London, United Kingdom: David & Charles.
Lyell, C., and J. W. Dawson. 1853. “On the Discovery of Some Fossil Reptilian Remains, and a Land-Shell in the Interior of an Erect Fossil-Tree in the Coal Measures of Nova Scotia, with Remarks on the Origin of Coal-Fields, and the Time Required for their Formation.” American Journal of Science 16: 33–41.
MacFadyen, Colin and John Gordon. 2006. Glasgow and Ayrshire: A Landscape Fashioned by Geology. Battleby, Scotland: Scottish National Heritage.
MacGregor, Murray and John Walton. 1948. The Story of the Fossil Grove. Glasgow, United Kingdom: City of Glasgow Public Parks and Botanic Gardens Department.
MacGregor, Murray and John Walton. 1983. The Story of the Fossil Grove, 5th edition. Glasgow, United Kingdom: Glasgow District Council Department of Parks and Recreation.
McCallien, William John. 1938. Geology of Glasgow and District. London, United Kingdom: Blackie and Son Ltd.
McLean, A. C. 1973. “Excursion 1: Fossil Grove.” In Excursion Guide to the Geology of the Glasgow District, edited by B. J. Bluck, 29–32. Glasgow, United Kingdom: Geological Society of Glasgow.
Monaghan, Alison A. 2014. The Carboniferous Shales of the Midland Valley of Scotland: Geology and Resource Estimation. London, United Kingdom: British Geological Survey for Department of Energy and Climate Change. https://www.ogauthority.co.uk/media/2765/bgs_decc_mvs_2014_main_report.pdf.
Neilson, J. 1901. “The ‘Fossil Grove’ in the Victoria Park, Whiteinch.” Excursion [#10] to Fossil Grove, Whiteinch, Glasgow Meeting [of the British Association for the Advancement of Science]. London, United Kingdom: The British Association for the Advancement of Science.
Oard, Michael J. and Hank Giesecke. 2007. “Polystrate Fossils Require Rapid Deposition.” Creation Research Society Quarterly 43, no. 3 (March): 232–240.
Owen, A., N. Everett, S. Leishman, and A. Gunning 2007. “Fossil Grove to be an Undercover RIGS.” Earth Heritage: The Geological and Landscape Conservation Magazine 29: 22–23. http://www.earthheritage.org.uk/wp/wp-content/uploads/2018/03/eh291-8.pdf.
Read, W. A., M. A. E. Browne, D. Stephenson, and B. G. J. Upton. 2002. “Carboniferous.” In The Geology of Scotland, 4th edition, edited by Nigel H. Trewin, 251–299. London, United Kingdom: The Geological Society.
Scheven, J. 1981. “Floating Forests on Firm Grounds: Advances in Carboniferous Research.” Journal of the Biblical Creation Society 3, no. 9: 36–43.
Taylor, Paul D. and Olev Vinn. 2006. “Convergence in Small Spiral Worm Tubes (‘Spirorbis’) and its Palaeoenvironmental Implications.” Journal of the Geological Society 163, no. 2 (March): 225–228.
Wieland, Carl. 1995. “Forests that Grew on Water: Startling Hard Facts from Coal Uproot the ‘Millions of Years’ Idea.” Creation 18, no. 1 (December): 20–24.
Wise, K. P. 2003. “The Pre-Flood Floating Forest: A Study in Paleontological Pattern Recognition.” In Proceedings of the Fifth International Conference on Creationism, edited by Robert L. Ivey Jr., 371–381. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Young, John and D. Corse Glen. 1888. “Notes on a Section of Carboniferous Strata, Containing Erect Stems of Fossil Trees and Beds of Intrusive Dolerite, in the Old Whinstone Quarry, Victoria Park, Lower Balshagray, near Whiteinch and Partick.” Transactions of the Geological Society of Glasgow 8, no. 2: 227–235.














